

An opioid agonist that does not induce mu-opioid receptor--arrestin interactions or receptor internalization
- PMID: 17090705
- PMCID: PMC3926195
- DOI: 10.1124/mol.106.028258
An opioid agonist that does not induce mu-opioid receptor--arrestin interactions or receptor internalization
Abstract
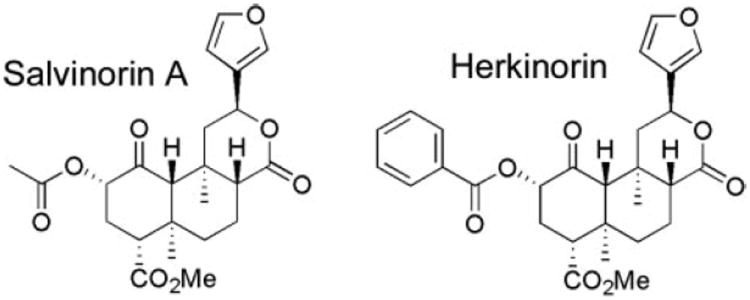
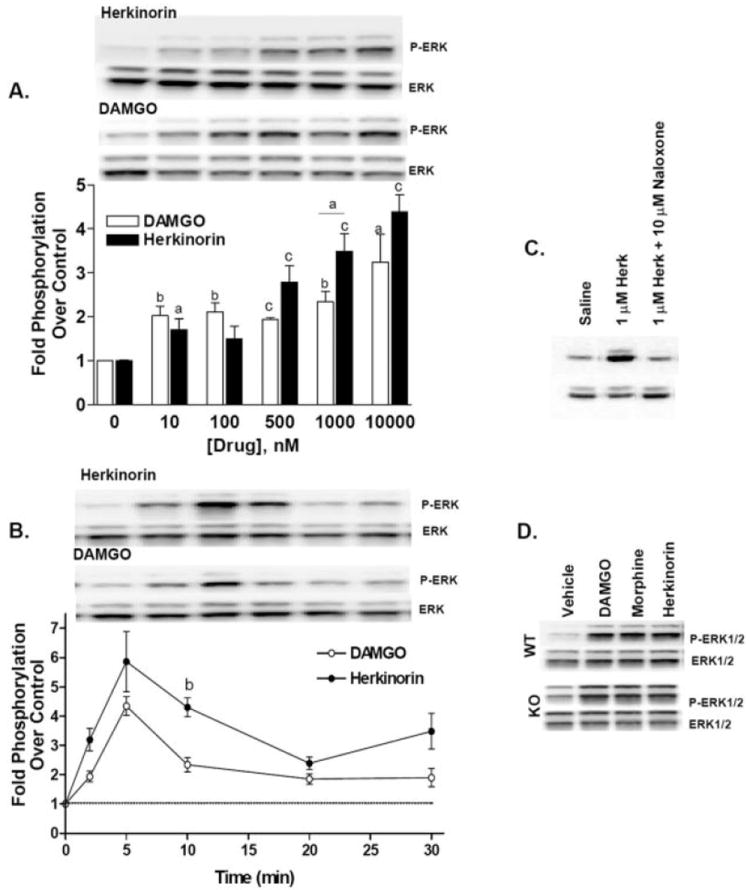
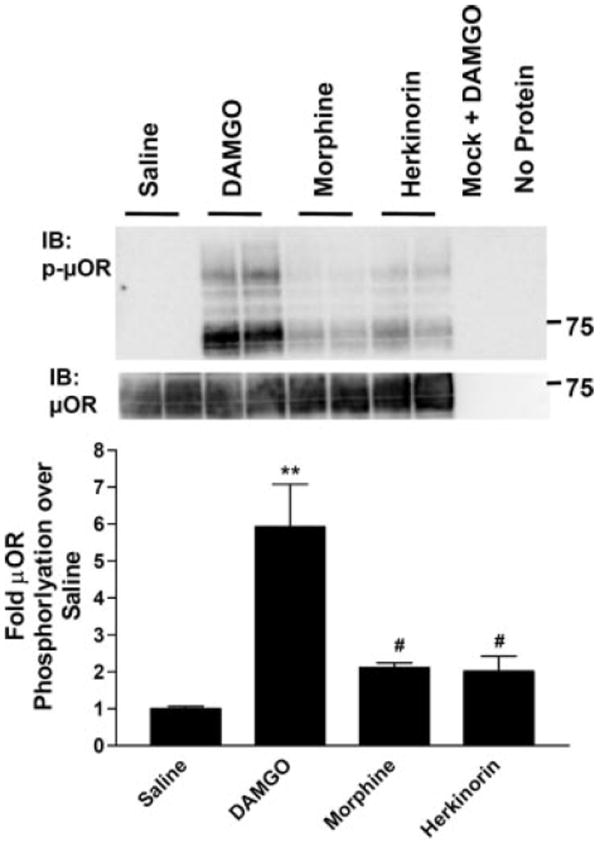
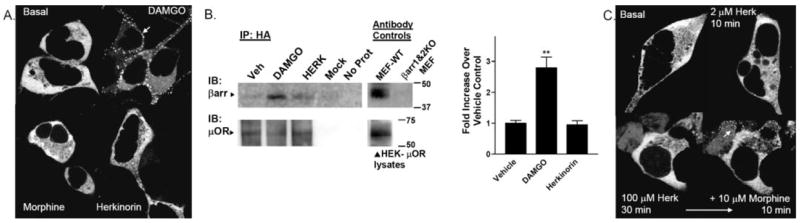
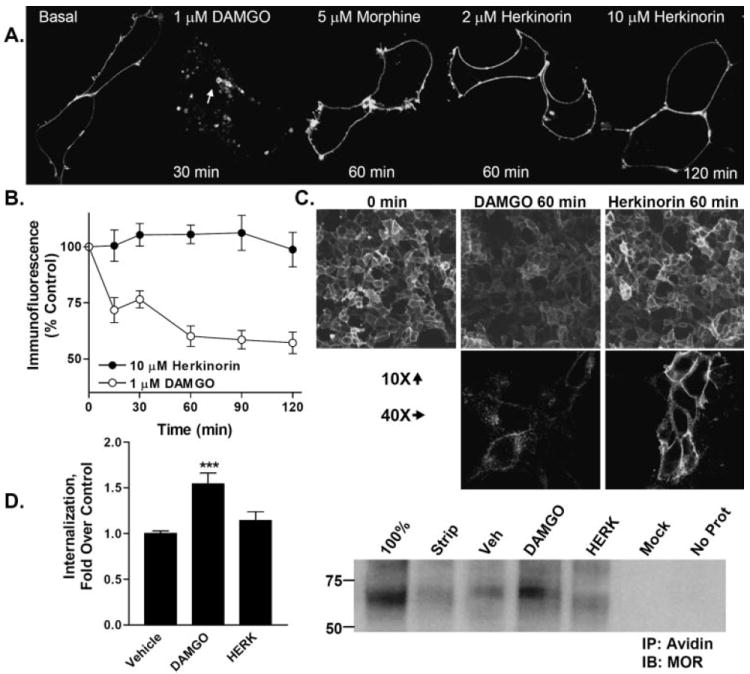
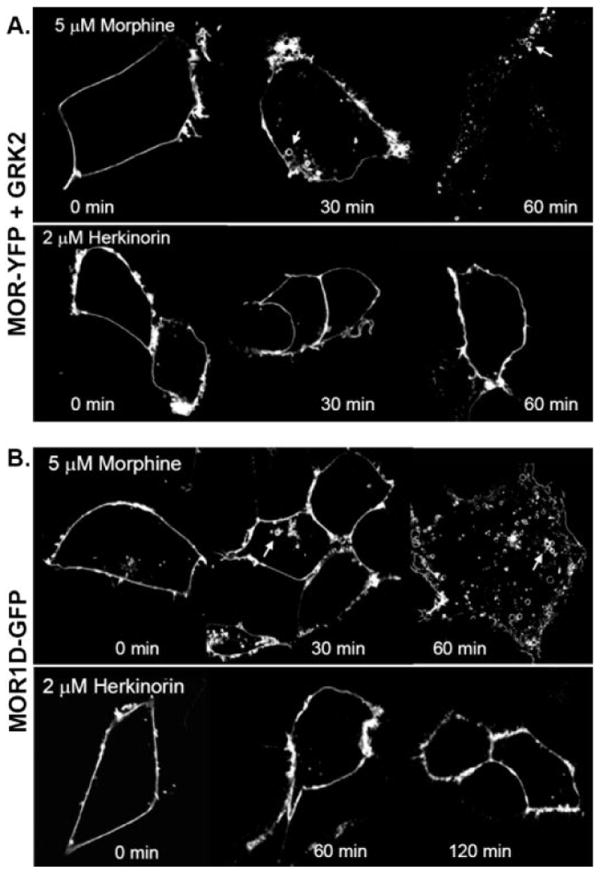
G protein-coupled receptor desensitization and trafficking are important regulators of opioid receptor signaling that can dictate overall drug responsiveness in vivo. Furthermore, different mu-opioid receptor (muOR) ligands can lead to varying degrees of receptor regulation, presumably because of distinct structural conformations conferred by agonist binding. For example, morphine binding produces a muOR with low affinity for beta-arrestin proteins and limited receptor internalization, whereas enkephalin analogs promote robust trafficking of both beta-arrestins and the receptors. Here, we evaluate muOR trafficking in response to activation by a novel mu-selective agonist derived from the naturally occurring plant product, salvinorin A. It is interesting that this compound, termed herkinorin, does not promote the recruitment of beta-arrestin-2 to the muOR and does not lead to receptor internalization. Moreover, whereas G protein-coupled receptor kinase overexpression can promote morphine-induced beta-arrestin interactions and muOR internalization, such manipulations do not promote herkinorin-induced trafficking. Studies in mice have shown that beta-arrestin-2 plays an important role in the development of morphine-induced tolerance, constipation, and respiratory depression. Therefore, drugs that can activate the receptor without recruiting the arrestins may be a promising step in the development of opiate analgesics that distinguish between agonist activity and receptor regulation and may ultimately lead to therapeutics designed to provide pain relief without the adverse side effects normally associated with the opiate narcotics.
Figures
References
-
- Alvarez V, Arttamangkul S, Williams JT. A RAVE about opioid withdrawal. Neuron. 2001;32:761–763. - PubMed
-
- Bohn LM, Dykstra LA, Lefkowitz RJ, Caron MG, Barak LS. Relative opioid efficacy is determined by the complements of the G protein-coupled receptor desensitization machinery. Mol Pharmacol. 2004a;66:106–112. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
