

The dependence of enzyme activity on temperature: determination and validation of parameters
- PMID: 17092210
- PMCID: PMC1798444
- DOI: 10.1042/BJ20061143
The dependence of enzyme activity on temperature: determination and validation of parameters
Erratum in
- Biochem J. 2007 May1:403(3):615
Abstract
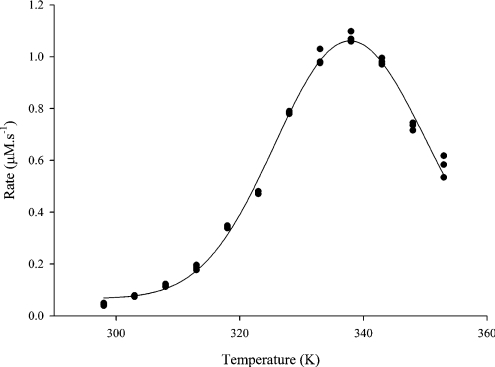
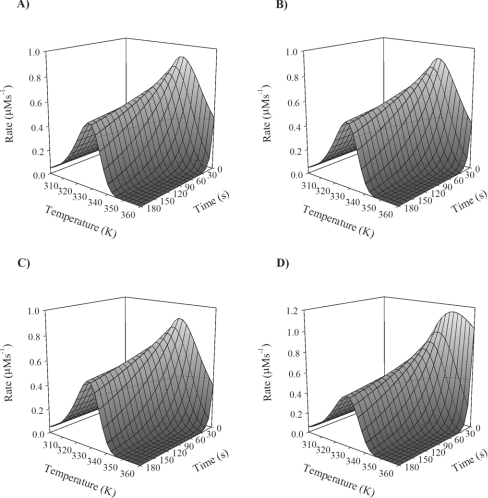
Traditionally, the dependence of enzyme activity on temperature has been described by a model consisting of two processes: the catalytic reaction defined by DeltaG(Dagger)(cat), and irreversible inactivation defined by DeltaG(Dagger)(inact). However, such a model does not account for the observed temperature-dependent behaviour of enzymes, and a new model has been developed and validated. This model (the Equilibrium Model) describes a new mechanism by which enzymes lose activity at high temperatures, by including an inactive form of the enzyme (E(inact)) that is in reversible equilibrium with the active form (E(act)); it is the inactive form that undergoes irreversible thermal inactivation to the thermally denatured state. This equilibrium is described by an equilibrium constant whose temperature-dependence is characterized in terms of the enthalpy of the equilibrium, DeltaH(eq), and a new thermal parameter, T(eq), which is the temperature at which the concentrations of E(act) and E(inact) are equal; T(eq) may therefore be regarded as the thermal equivalent of K(m). Characterization of an enzyme with respect to its temperature-dependent behaviour must therefore include a determination of these intrinsic properties. The Equilibrium Model has major implications for enzymology, biotechnology and understanding the evolution of enzymes. The present study presents a new direct data-fitting method based on fitting progress curves directly to the Equilibrium Model, and assesses the robustness of this procedure and the effect of assay data on the accurate determination of T(eq) and its associated parameters. It also describes simpler experimental methods for their determination than have been previously available, including those required for the application of the Equilibrium Model to non-ideal enzyme reactions.
Figures

Similar articles
-
New parameters controlling the effect of temperature on enzyme activity.Biochem Soc Trans. 2007 Dec;35(Pt 6):1543-6. doi: 10.1042/BST0351543. Biochem Soc Trans. 2007. PMID: 18031263 Review.
-
Eurythermalism and the temperature dependence of enzyme activity.FASEB J. 2007 Jun;21(8):1934-41. doi: 10.1096/fj.06-7265com. Epub 2007 Mar 6. FASEB J. 2007. PMID: 17341686
-
Determination of enzyme thermal parameters for rational enzyme engineering and environmental/evolutionary studies.Methods Mol Biol. 2013;996:219-30. doi: 10.1007/978-1-62703-354-1_13. Methods Mol Biol. 2013. PMID: 23504427
-
The molecular basis of the effect of temperature on enzyme activity.Biochem J. 2009 Dec 23;425(2):353-60. doi: 10.1042/BJ20091254. Biochem J. 2009. PMID: 19849667
-
A new understanding of how temperature affects the catalytic activity of enzymes.Trends Biochem Sci. 2010 Oct;35(10):584-91. doi: 10.1016/j.tibs.2010.05.001. Epub 2010 Jun 16. Trends Biochem Sci. 2010. PMID: 20554446 Review.
Cited by
-
Intracellular temperature mapping with fluorescence-assisted photoacoustic-thermometry.Appl Phys Lett. 2013 May 13;102(19):193705. doi: 10.1063/1.4807140. Epub 2013 May 16. Appl Phys Lett. 2013. PMID: 23825801 Free PMC article.
-
Acetylcholinesterase- and Butyrylcholinesterase-Based Biosensors for the Detection of Quaternary Ammonium Biocides in Food Industry.Foods. 2023 Dec 30;13(1):133. doi: 10.3390/foods13010133. Foods. 2023. PMID: 38201162 Free PMC article.
-
Macroalgal Proteins: A Review.Foods. 2022 Feb 16;11(4):571. doi: 10.3390/foods11040571. Foods. 2022. PMID: 35206049 Free PMC article. Review.
-
Continuous-Flow High-Pressure Homogenization of Blueberry Juice Enhances Anthocyanin and Ascorbic Acid Stability during Cold Storage.J Agric Food Chem. 2024 May 22;72(20):11629-11639. doi: 10.1021/acs.jafc.4c01289. Epub 2024 May 13. J Agric Food Chem. 2024. PMID: 38739462 Free PMC article.
-
Characterization and evolution of a metagenome-derived lipase towards enhanced enzyme activity and thermostability.Mol Cell Biochem. 2013 Jan;373(1-2):149-59. doi: 10.1007/s11010-012-1483-8. Epub 2012 Oct 27. Mol Cell Biochem. 2013. PMID: 23104399
References
-
- Daniel R. M., Danson M. J., Eisenthal R. The temperature optima of enzymes: a new perspective on an old phenomenon. Trends Biochem. Sci. 2001;26:223–225. - PubMed
-
- Peterson M. E., Eisenthal R., Danson M. J., Spence A., Daniel R. M. A new, intrinsic, thermal parameter for enzymes reveals true temperature optima. J. Biol. Chem. 2004;279:20717–20722. - PubMed
-
- Eisenthal R., Peterson M. E., Daniel R. M., Danson M. J. The thermal behaviour of enzymes: implications for biotechnology. Trends Biotechnol. 2006;24:289–292. - PubMed
-
- Daniel R. M., Danson M. J. Assaying activity and assessing thermostability of hyperthermophilic enzymes. Methods Enzymol. 2001;334:283–293. - PubMed
-
- Tipton K. F. Principles of enzymes assays and kinetic studies. In: Eisenthal R., Danson M. J., editors. Enzyme Assays, a Practical Approach. Oxford: Oxford University Press; 1992. pp. 1–58.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous