

Viscoelastic properties of individual glial cells and neurons in the CNS
- PMID: 17093050
- PMCID: PMC1693820
- DOI: 10.1073/pnas.0606150103
Viscoelastic properties of individual glial cells and neurons in the CNS
Abstract
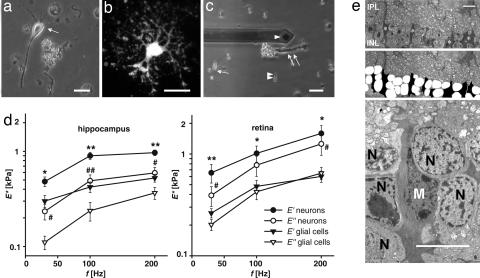
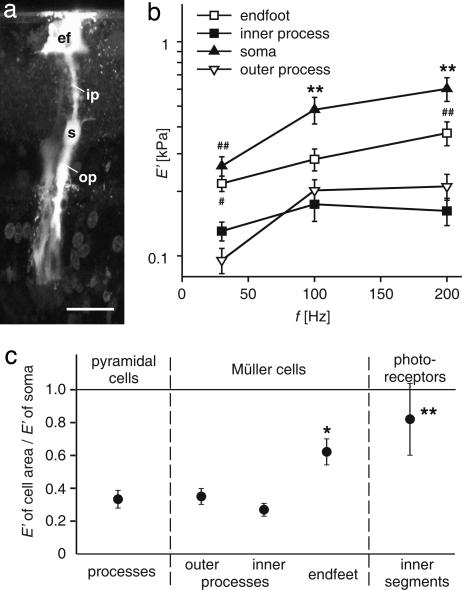
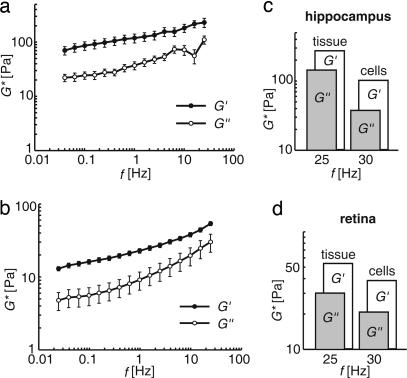
One hundred fifty years ago glial cells were discovered as a second, non-neuronal, cell type in the central nervous system. To ascribe a function to these new, enigmatic cells, it was suggested that they either glue the neurons together (the Greek word "gammalambdaiotaalpha" means "glue") or provide a robust scaffold for them ("support cells"). Although both speculations are still widely accepted, they would actually require quite different mechanical cell properties, and neither one has ever been confirmed experimentally. We investigated the biomechanics of CNS tissue and acutely isolated individual neurons and glial cells from mammalian brain (hippocampus) and retina. Scanning force microscopy, bulk rheology, and optically induced deformation were used to determine their viscoelastic characteristics. We found that (i) in all CNS cells the elastic behavior dominates over the viscous behavior, (ii) in distinct cell compartments, such as soma and cell processes, the mechanical properties differ, most likely because of the unequal local distribution of cell organelles, (iii) in comparison to most other eukaryotic cells, both neurons and glial cells are very soft ("rubber elastic"), and (iv) intriguingly, glial cells are even softer than their neighboring neurons. Our results indicate that glial cells can neither serve as structural support cells (as they are too soft) nor as glue (because restoring forces are dominant) for neurons. Nevertheless, from a structural perspective they might act as soft, compliant embedding for neurons, protecting them in case of mechanical trauma, and also as a soft substrate required for neurite growth and facilitating neuronal plasticity.
Conflict of interest statement
The authors declare no conflict of interest.
Figures

References
-
- Virchow R. Gesammelte Abhandlungen zur Wissenschaftlichen Medicin. Frankfurt, Germany: von Meidinger Sohn; 1856.
-
- Schultze M. Zur Anatomie und Physiologie der Retina. Sohn, Bonn, Germany: von Max Cohen; 1866.
-
- Arbogast KB, Margulies SS. J Biomech. 1998;31:801–807. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
