

Cue-invariant networks for figure and background processing in human visual cortex
- PMID: 17093091
- PMCID: PMC2711040
- DOI: 10.1523/JNEUROSCI.2741-06.2006
Cue-invariant networks for figure and background processing in human visual cortex
Abstract
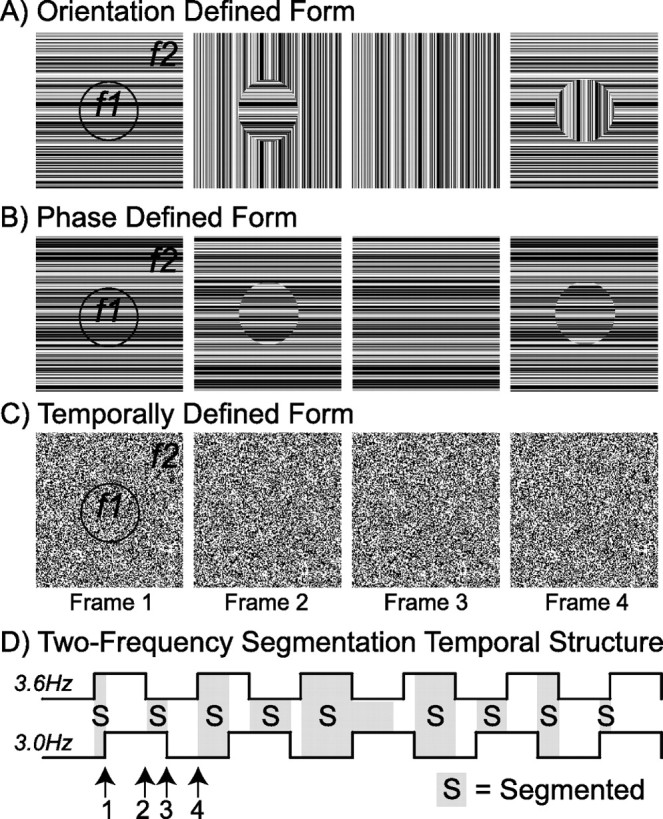
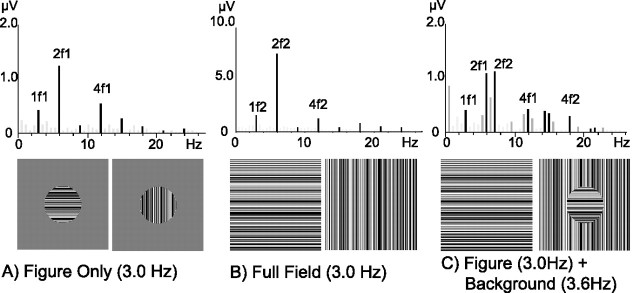
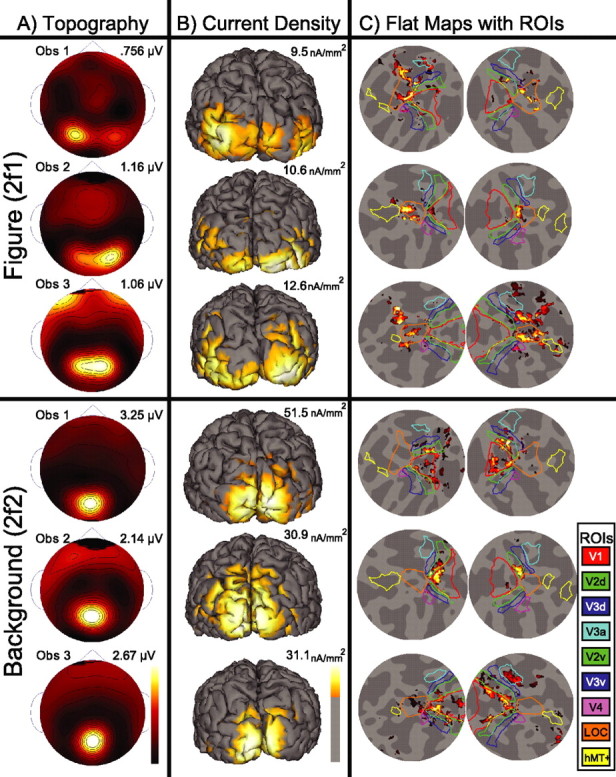
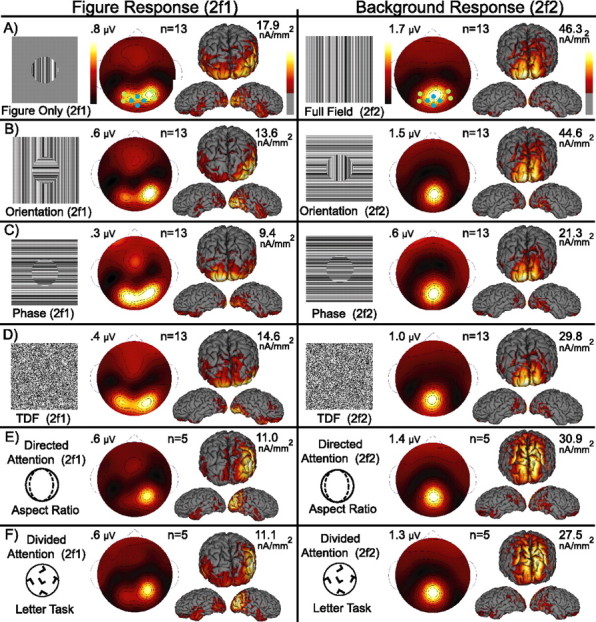
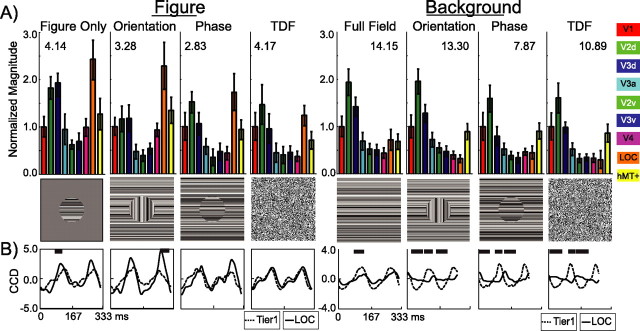
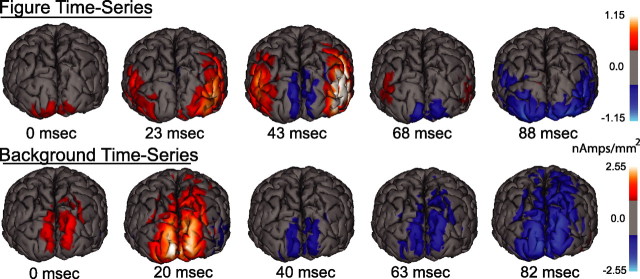
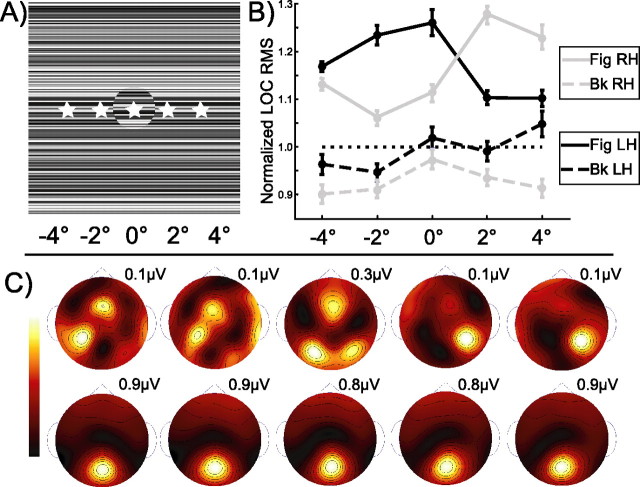
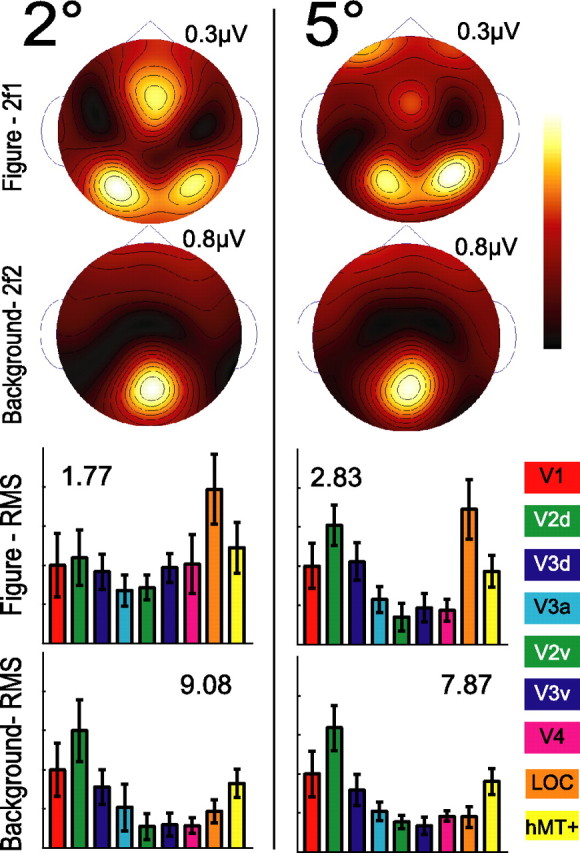
Lateral occipital cortical areas are involved in the perception of objects, but it is not clear how these areas interact with first tier visual areas. Using synthetic images portraying a simple texture-defined figure and an electrophysiological paradigm that allows us to monitor cortical responses to figure and background regions separately, we found distinct neuronal networks responsible for the processing of each region. The figure region of our displays was tagged with one temporal frequency (3.0 Hz) and the background region with another (3.6 Hz). Spectral analysis was used to separate the responses to the two regions during their simultaneous presentation. Distributed source reconstructions were made by using the minimum norm method, and cortical current density was measured in a set of visual areas defined on retinotopic and functional criteria with the use of functional magnetic resonance imaging. The results of the main experiments, combined with a set of control experiments, indicate that the figure region, but not the background, was routed preferentially to lateral cortex. A separate network extending from first tier through more dorsal areas responded preferentially to the background region. The figure-related responses were mostly invariant with respect to the texture types used to define the figure, did not depend on its spatial location or size, and mostly were unaffected by attentional instructions. Because of the emergent nature of a segmented figure in our displays, feedback from higher cortical areas is a likely candidate for the selection mechanism by which the figure region is routed to lateral occipital cortex.
Figures
References
-
- Bevington PR, Robinson DK. New York: McGraw-Hill; 1992. Data reduction and error analysis for the physical sciences.
-
- Blair RC, Karniski W. An alternative method for significance testing of waveform difference potentials. Psychophysiology. 1993;30:518–524. - PubMed
-
- Brewer AA, Liu J, Wade AR, Wandell BA. Visual field maps and stimulus selectivity in human ventral occipital cortex. Nat Neurosci. 2005;8:1102–1109. - PubMed
-
- Caputo G, Casco C. A visual evoked potential correlate of global figure-ground segmentation. Vision Res. 1999;39:1597–1610. - PubMed
-
- Casco C, Grieco A, Campana G, Corvino MP, Caputo G. Attention modulates psychophysical and electrophysiological response to visual texture segmentation in humans. Vision Res. 2005;45:2384–2396. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources