

Adenosine inhibits activity of hypocretin/orexin neurons by the A1 receptor in the lateral hypothalamus: a possible sleep-promoting effect
- PMID: 17093123
- PMCID: PMC1783688
- DOI: 10.1152/jn.00873.2006
Adenosine inhibits activity of hypocretin/orexin neurons by the A1 receptor in the lateral hypothalamus: a possible sleep-promoting effect
Abstract
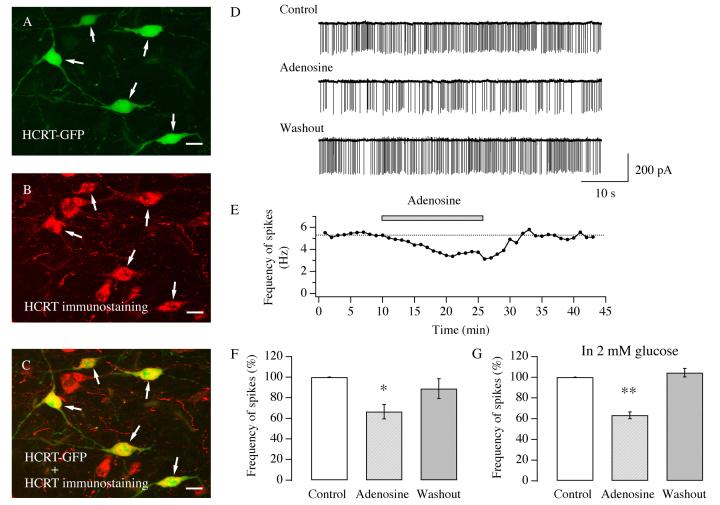
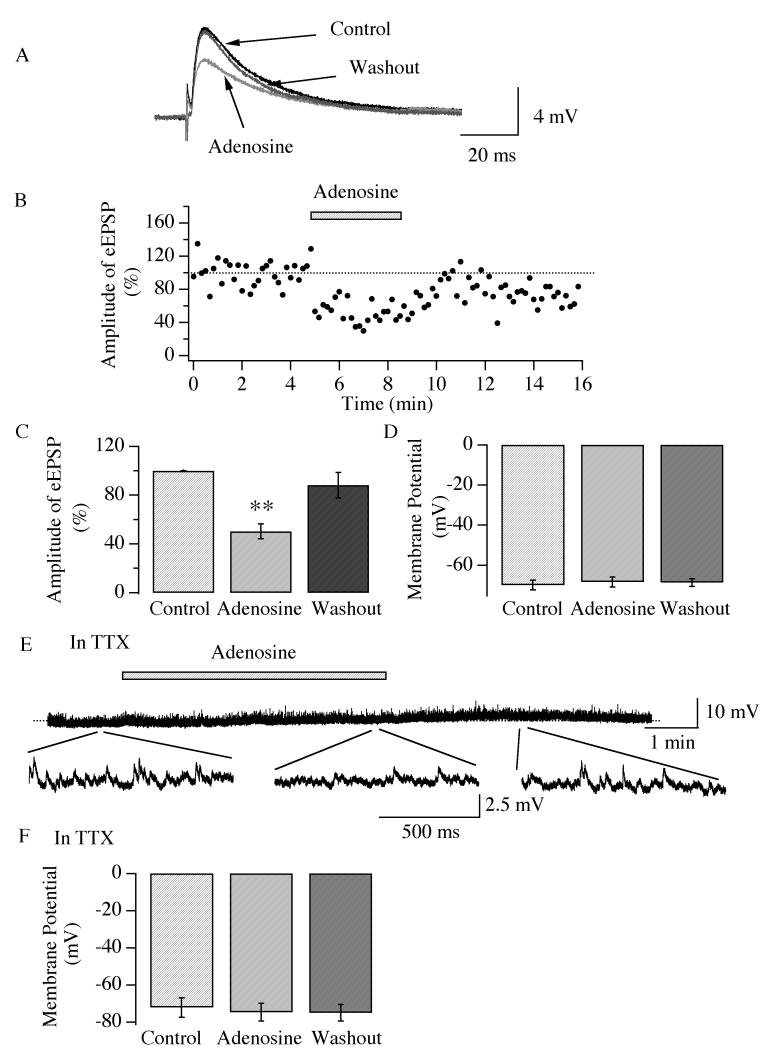
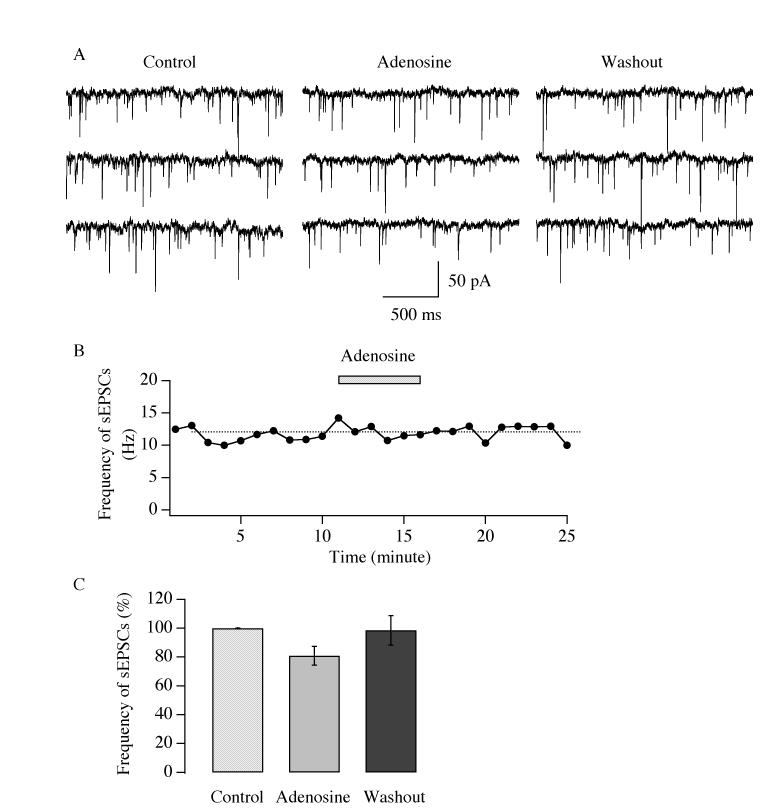
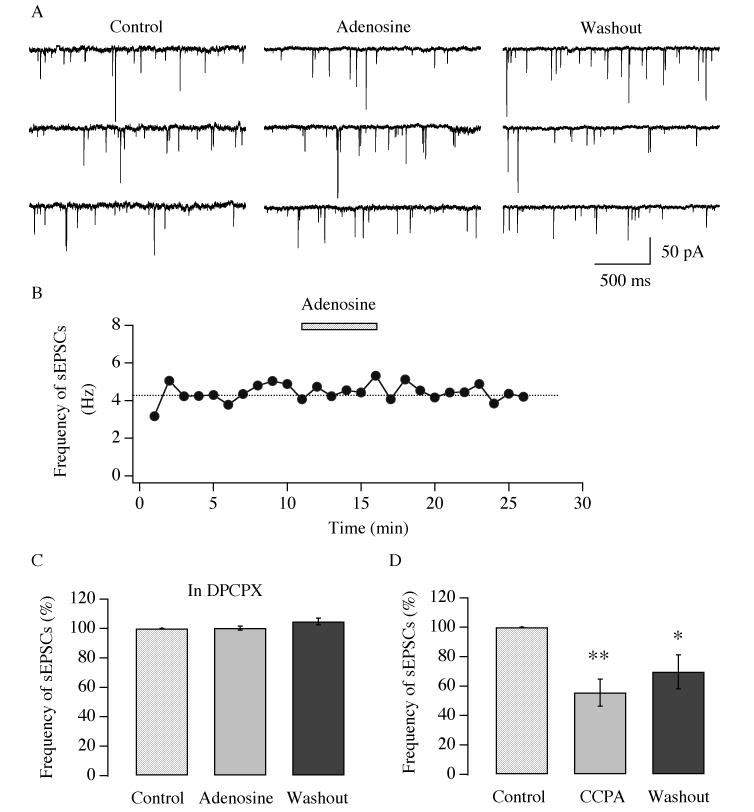
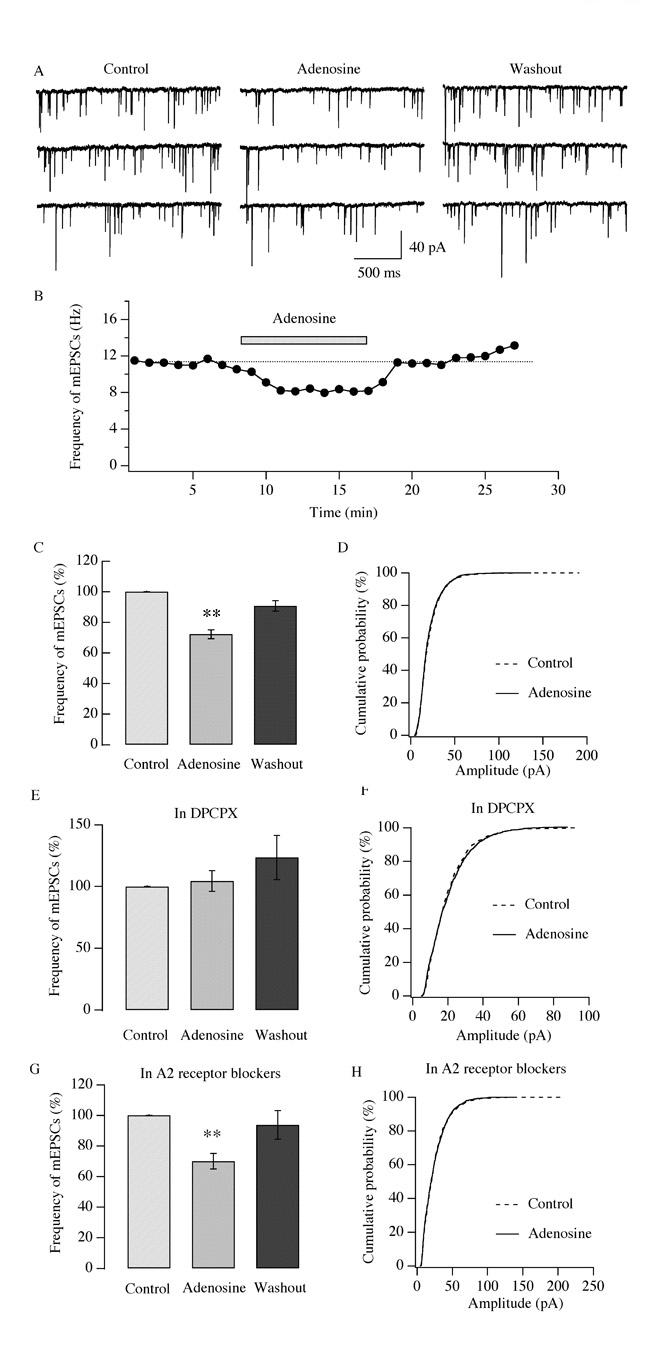
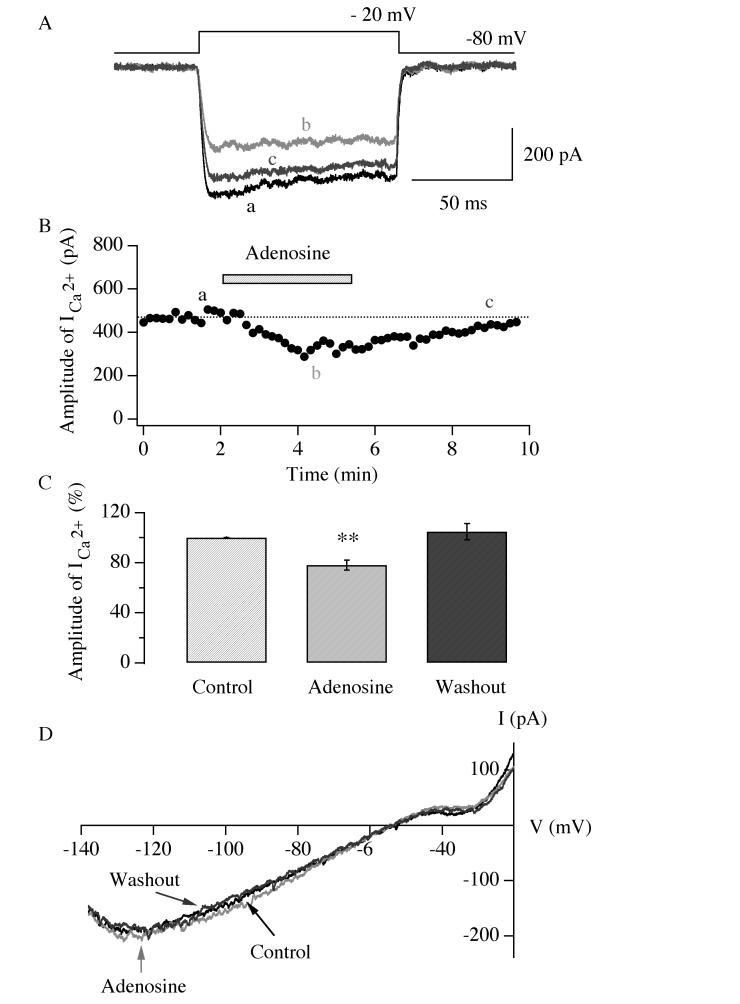
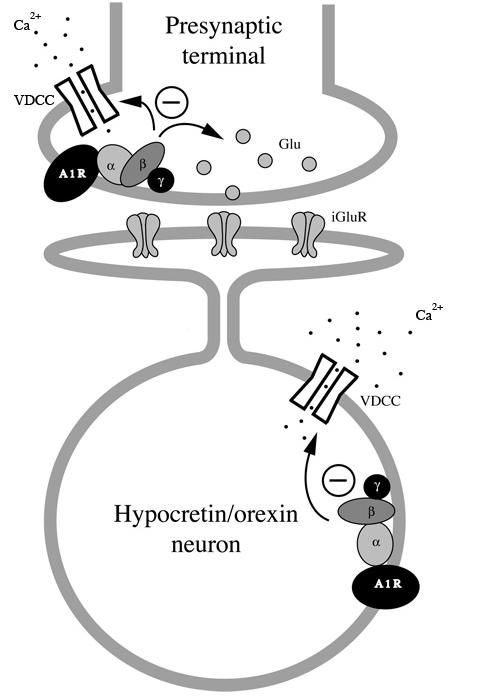
Neurons in the lateral hypothalamus (LH) that contain hypocretin/orexin have been established as important promoters of arousal. Deficiencies in the hypocretin/orexin system lead to narcolepsy. The inhibition of hypocretin/orexin neurons by sleep-promoting neurotransmitters has been suggested as one part of the sleep regulation machinery. Adenosine has been identified as a sleep promoter and its role in sleep regulation in the basal forebrain has been well documented. However, the effect of adenosine on arousal-promoting hypocretin/orexin neurons has not been addressed, despite recent evidence that immunocytochemical visualization of adenosine receptors was detected in these neurons. In this study, we examined the hypothesis that adenosine inhibits the activity of hypocretin/orexin neurons by using electrophysiological methods in brain slices from mice expressing green fluorescent protein in hypocretin/orexin neurons. We found that adenosine significantly attenuated the frequency of action potentials without a change in membrane potential in hypocretin/orexin neurons. The adenosine-mediated inhibition arises from depression of excitatory synaptic transmission to hypocretin/orexin neurons because adenosine depresses the amplitude of evoked excitatory postsynaptic potential and the frequency of spontaneous and miniature excitatory postsynaptic currents in these neurons. At the cell body of the hypocretin/orexin neurons, adenosine inhibits voltage-dependent calcium currents without the induction of GIRK current. The inhibitory effect of adenosine is dose dependent, pertussis toxin sensitive, and mediated by A1 receptors. In summary, our data suggest that in addition to its effect in the basal forebrain, adenosine exerts its sleep-promoting effect in the LH by inhibition of hypocretin/orexin neurons.
Figures


References
-
- Abel T, Nguyen PV, Barad M, Deuel TA, Kandel ER, Bourthouladze R. Genetic demonstration of a role for PKA in the late phase of LTP and in hippocampus-based longterm memory. Cell. 1997;88:615–626. - PubMed
-
- Arrigoni E, Chamberlin NL, Saper CB, McCarley RW. Adenosine inhibits basal forebrain cholinergic and noncholinergic neurons in vitro. Neuroscience. 2006;140:403–413. - PubMed
-
- Bading H, Ginty DD, Greenberg ME. Regulation of gene expression in hippocampal neurons by distinct calcium signaling pathways. Science. 1993;260:181–186. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources