

Biochemical and genetic analysis of the vaccinia virus d5 protein: Multimerization-dependent ATPase activity is required to support viral DNA replication
- PMID: 17093187
- PMCID: PMC1797480
- DOI: 10.1128/JVI.02217-06
Biochemical and genetic analysis of the vaccinia virus d5 protein: Multimerization-dependent ATPase activity is required to support viral DNA replication
Abstract
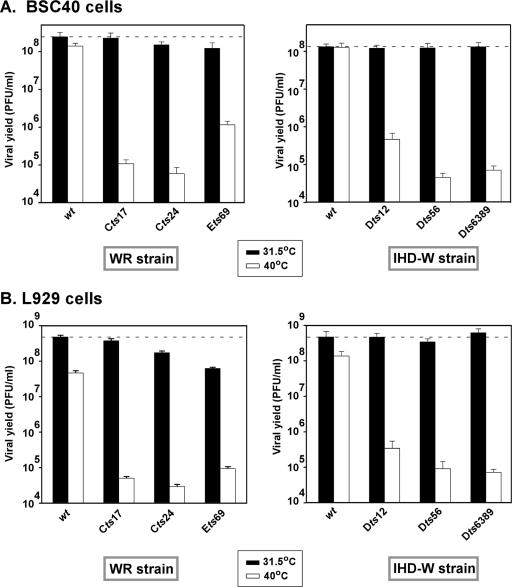
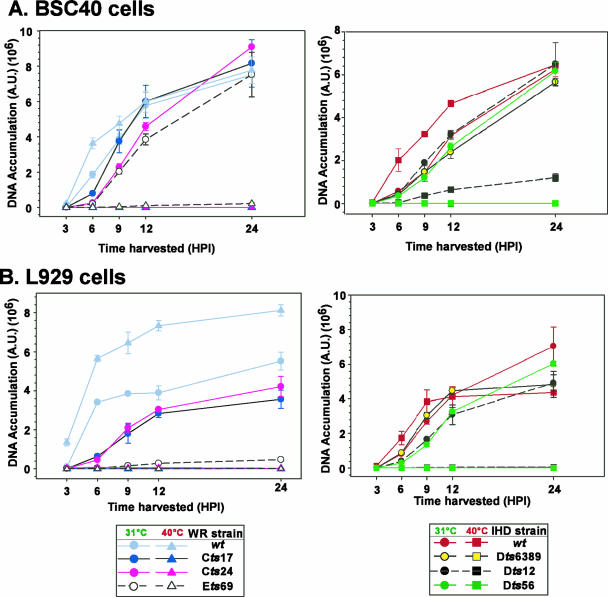
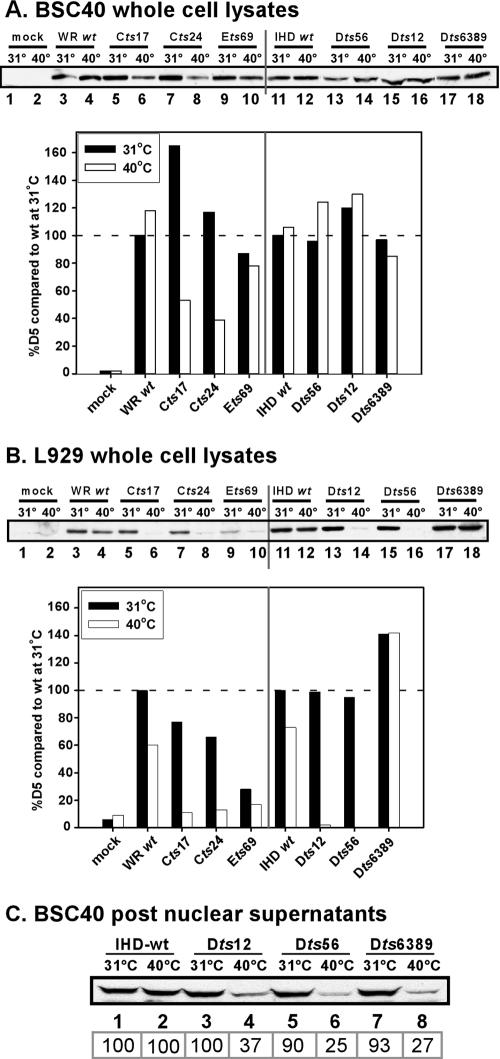
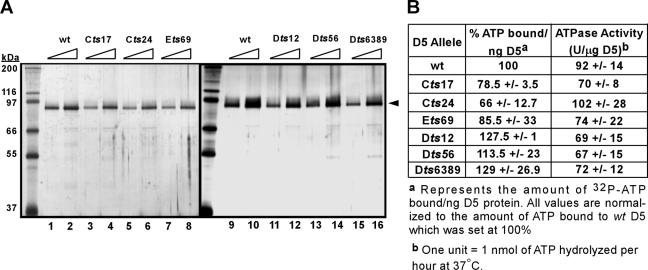
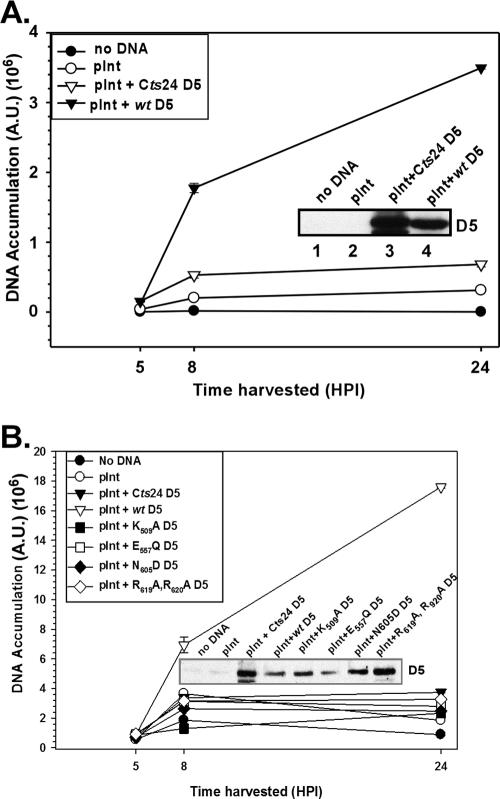
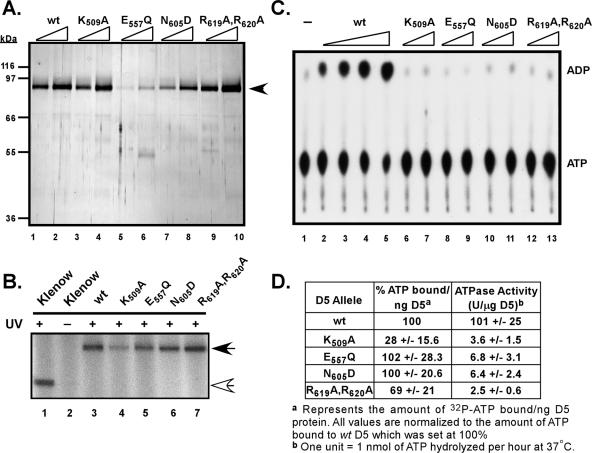
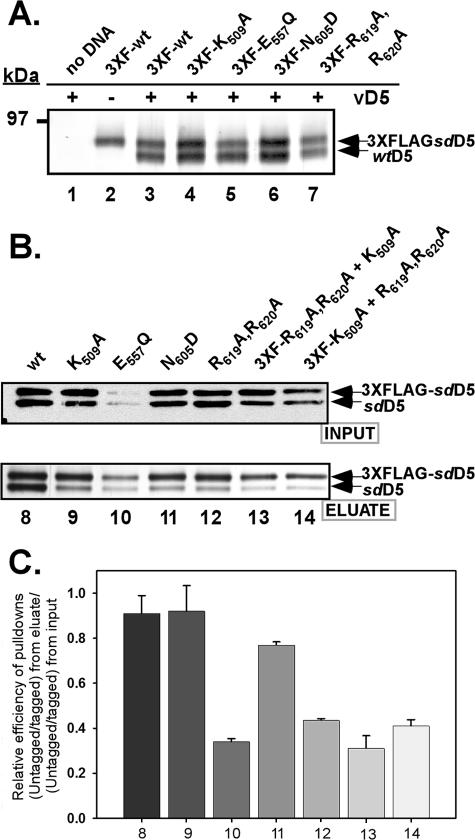
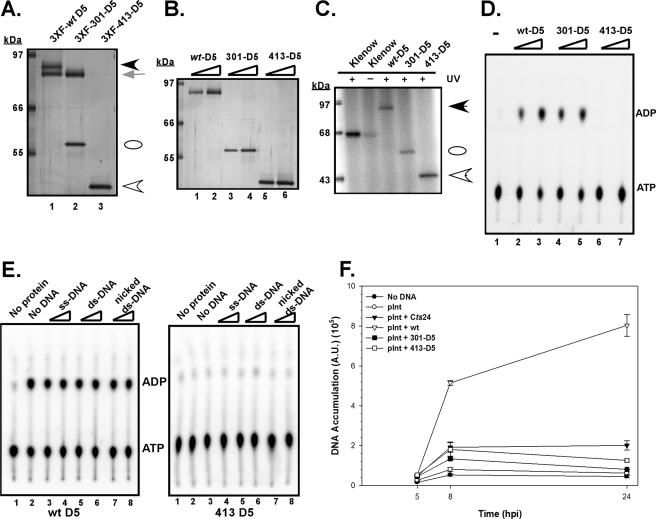
The vaccinia virus-encoded D5 protein is an essential ATPase involved in viral DNA replication. We have expanded the genotypic and phenotypic analysis of six temperature-sensitive (ts) D5 mutants (Cts17, Cts24, Ets69, Dts6389 [also referred to as Dts38], Dts12, and Dts56) and shown that at nonpermissive temperature all of the tsD5 viruses exhibit a dramatic reduction in DNA synthesis and virus production. For Cts17 and Cts24, this restriction reflects the thermolability of the D5 proteins. The Dts6389, Dts12, and Dts56 D5 proteins become insoluble at 39.7 degrees C, while the Ets69 D5 protein remains stable and soluble and retains the ability to oligomerize and hydrolyze ATP when synthesized at 39.7 degrees C. To investigate which structural features of D5 are important for its biological and biochemical activities, we generated targeted mutations in invariant residues positioned within conserved domains found within D5. Using a transient complementation assay that assessed the ability of D5 variants to sustain ongoing DNA synthesis during nonpermissive Cts24 infections, only a wtD5 allele supported DNA synthesis. Alleles of D5 containing targeted mutations within the Walker A or B domains, the superfamily III helicase motif C, or the AAA+ motif lacked biological competency. Furthermore, purified preparations of these variant proteins revealed that they all were defective in ATP hydrolysis. Multimerization of D5 appeared to be a prerequisite for enzymatic activity and required the Walker B domain, the AAA+ motif, and a region located upstream of the catalytic core. Finally, although multimerization and enzymatic activity are necessary for the biological competence of D5, they are not sufficient.
Figures

References
-
- Challberg, M. D., and P. T. Englund. 1979. Purification and properties of the deoxyribonucleic acid polymerase induced by vaccinia virus. J. Biol. Chem. 254:7812-7819. - PubMed
-
- Challberg, M. D., and P. T. Englund. 1979. The effect of template secondary structure on vaccinia DNA polymerase. J. Biol. Chem. 254:7820-7826. - PubMed
-
- Condit, R. C., A. Motyczka, and G. Spizz. 1983. Isolation, characterization, and physical mapping of temperature-sensitive mutants of vaccinia virus. Virology 128:429-443. - PubMed
-
- Dales, S., V. Milovanovitch, B. G. Pogo, S. B. Weintraub, T. Huima, S. Wilton, and G. McFadden. 1978. Biogenesis of vaccinia: isolation of conditional lethal mutants and electron microscopic characterization of their phenotypically expressed defects. Virology 84:403-428. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
