

Characterization of the preprotein and amino acid transporter gene family in Arabidopsis
- PMID: 17098851
- PMCID: PMC1761978
- DOI: 10.1104/pp.106.090688
Characterization of the preprotein and amino acid transporter gene family in Arabidopsis
Abstract
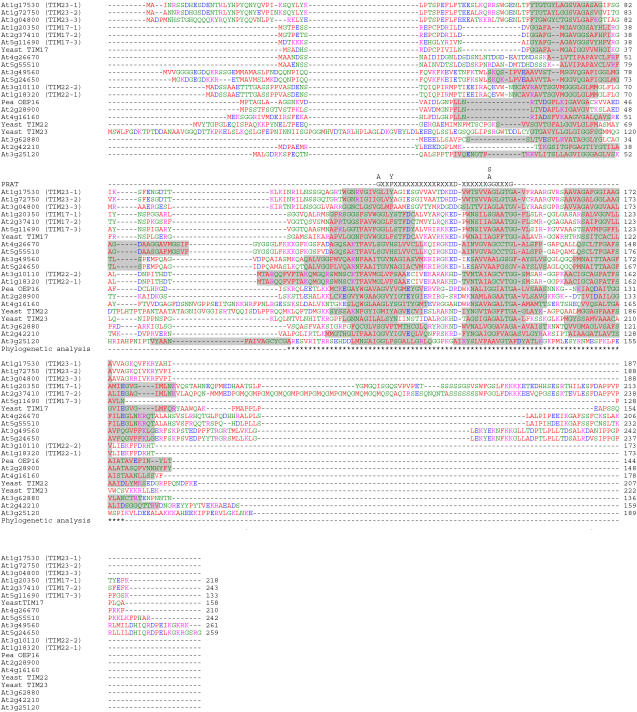
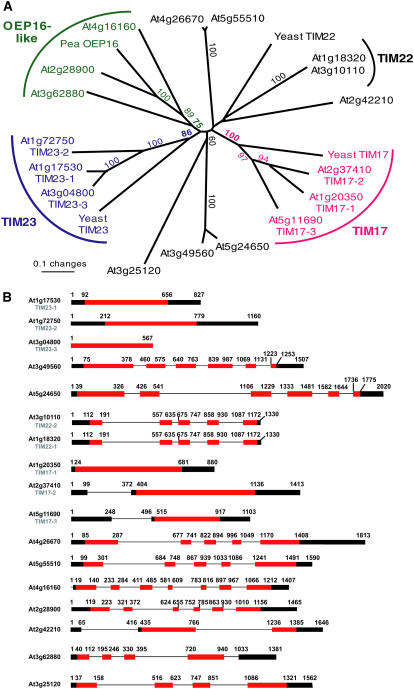
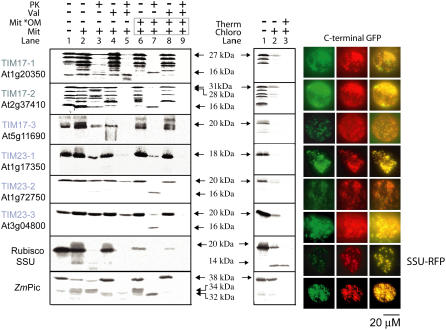
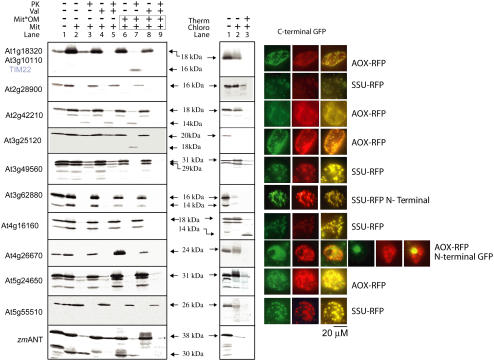
Seventeen loci encode proteins of the preprotein and amino acid transporter family in Arabidopsis (Arabidopsis thaliana). Some of these genes have arisen from recent duplications and are not in annotated duplicated regions of the Arabidopsis genome. In comparison to a number of other eukaryotic organisms, this family of proteins has greatly expanded in plants, with 24 loci in rice (Oryza sativa). Most of the Arabidopsis and rice genes are orthologous, indicating expansion of this family before monocot and dicot divergence. In vitro protein uptake assays, in vivo green fluorescent protein tagging, and immunological analyses of selected proteins determined either mitochondrial or plastidic localization for 10 and six proteins, respectively. The protein encoded by At5g24650 is targeted to both mitochondria and chloroplasts and, to our knowledge, is the first membrane protein reported to be targeted to mitochondria and chloroplasts. Three genes encoded translocase of the inner mitochondrial membrane (TIM)17-like proteins, three TIM23-like proteins, and three outer envelope protein16-like proteins in Arabidopsis. The identity of Arabidopsis TIM22-like proteins is most likely a protein encoded by At3g10110/At1g18320, based on phylogenetic analysis, subcellular localization, and complementation of a yeast (Saccharomyces cerevisiae) mutant and coexpression analysis. The lack of a preprotein and amino acid transporter domain in some proteins, localization in mitochondria, plastids, or both, variation in gene structure, and the differences in expression profiles indicate that the function of this family has diverged in plants beyond roles in protein translocation.
Figures


References
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
