

MAIZEWALL. Database and developmental gene expression profiling of cell wall biosynthesis and assembly in maize
- PMID: 17098859
- PMCID: PMC1761967
- DOI: 10.1104/pp.106.086405
MAIZEWALL. Database and developmental gene expression profiling of cell wall biosynthesis and assembly in maize
Abstract
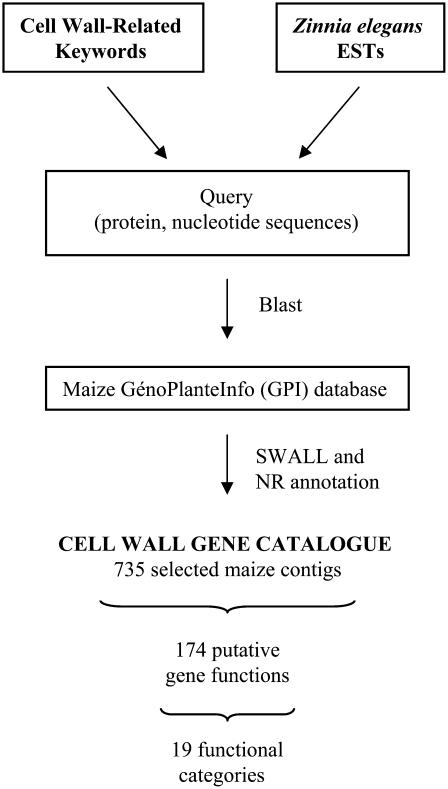
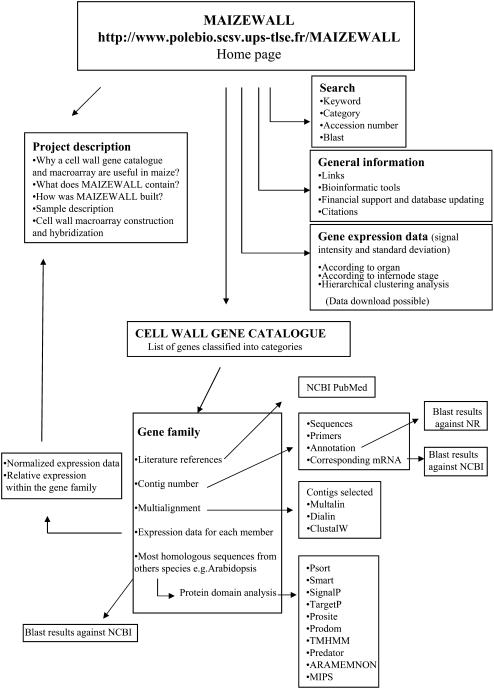
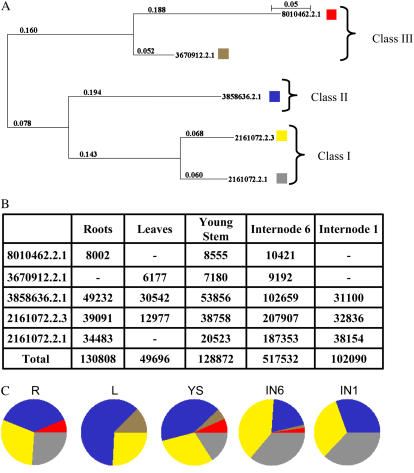
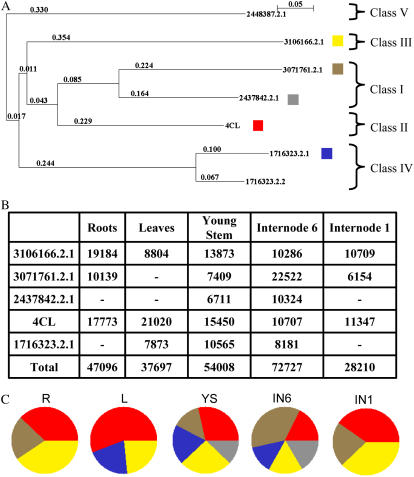
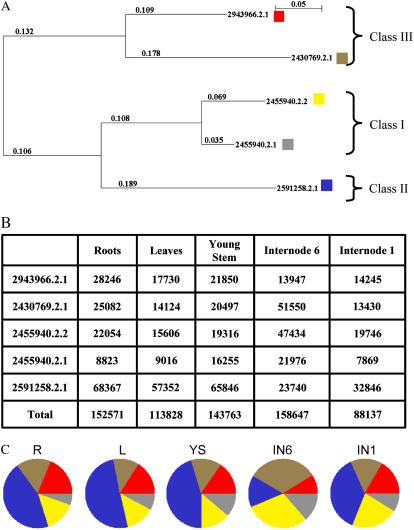
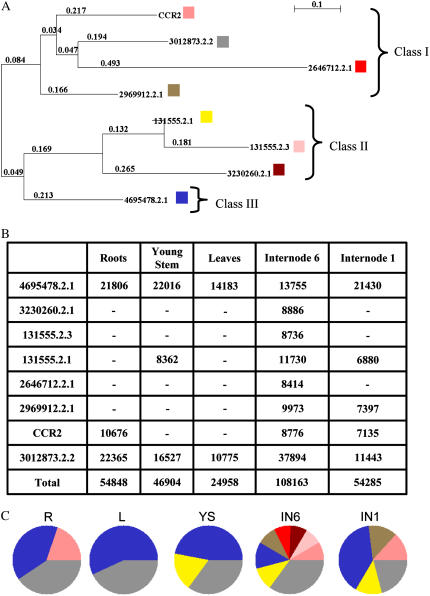
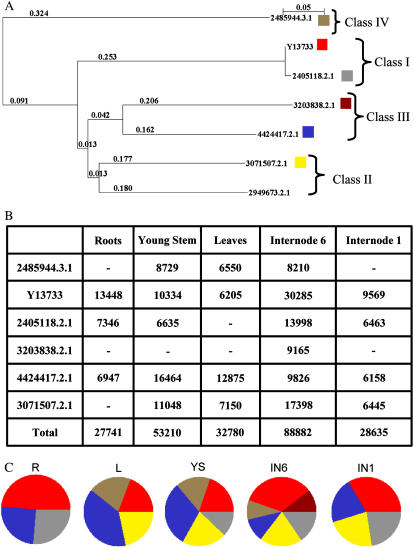
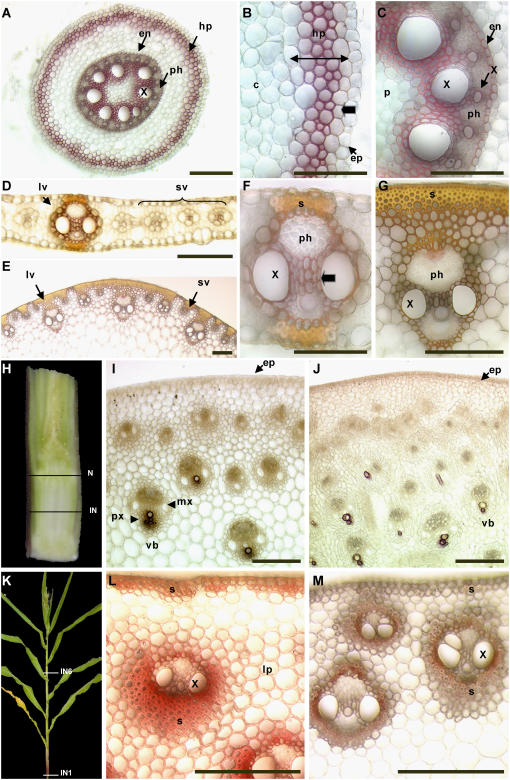
An extensive search for maize (Zea mays) genes involved in cell wall biosynthesis and assembly has been performed and 735 sequences have been centralized in a database, MAIZEWALL (http://www.polebio.scsv.ups-tlse.fr/MAIZEWALL). MAIZEWALL contains a bioinformatic analysis for each entry and gene expression data that are accessible via a user-friendly interface. A maize cell wall macroarray composed of a gene-specific tag for each entry was also constructed to monitor global cell wall-related gene expression in different organs and during internode development. By using this macroarray, we identified sets of genes that exhibit organ and internode-stage preferential expression profiles. These data provide a comprehensive fingerprint of cell wall-related gene expression throughout the maize plant. Moreover, an in-depth examination of genes involved in lignin biosynthesis coupled to biochemical and cytological data from different organs and stages of internode development has also been undertaken. These results allow us to trace spatially and developmentally regulated, putative preferential routes of monolignol biosynthesis involving specific gene family members and suggest that, although all of the gene families of the currently accepted monolignol biosynthetic pathway are conserved in maize, there are subtle differences in family size and a high degree of complexity in spatial expression patterns. These differences are in keeping with the diversity of lignified cell types throughout the maize plant.
Figures
Similar articles
-
Expression of cell wall related genes in basal and ear internodes of silking brown-midrib-3, caffeic acid O-methyltransferase (COMT) down-regulated, and normal maize plants.BMC Plant Biol. 2008 Jun 26;8:71. doi: 10.1186/1471-2229-8-71. BMC Plant Biol. 2008. PMID: 18582385 Free PMC article.
-
Genome-Wide Identification and Characterization of Lignin Synthesis Genes in Maize.Int J Mol Sci. 2024 Jun 18;25(12):6710. doi: 10.3390/ijms25126710. Int J Mol Sci. 2024. PMID: 38928419 Free PMC article.
-
Expression profiles of cell-wall related genes vary broadly between two common maize inbreds during stem development.BMC Genomics. 2019 Oct 29;20(1):785. doi: 10.1186/s12864-019-6117-z. BMC Genomics. 2019. PMID: 31664907 Free PMC article.
-
Trends in lignin modification: a comprehensive analysis of the effects of genetic manipulations/mutations on lignification and vascular integrity.Phytochemistry. 2002 Oct;61(3):221-94. doi: 10.1016/s0031-9422(02)00211-x. Phytochemistry. 2002. PMID: 12359514 Review.
-
Regulation of cell wall biosynthesis.Curr Opin Plant Biol. 2007 Dec;10(6):564-72. doi: 10.1016/j.pbi.2007.09.001. Epub 2007 Oct 22. Curr Opin Plant Biol. 2007. PMID: 17950657 Review.
Cited by
-
Diversity and linkage disequilibrium features in a composite public/private dent maize panel: consequences for association genetics as evaluated from a case study using flowering time.Theor Appl Genet. 2012 Aug;125(4):731-47. doi: 10.1007/s00122-012-1866-y. Epub 2012 May 24. Theor Appl Genet. 2012. PMID: 22622520
-
Proteomic analysis of embryo development in rice (Oryza sativa).Planta. 2012 Apr;235(4):687-701. doi: 10.1007/s00425-011-1535-4. Epub 2011 Oct 20. Planta. 2012. PMID: 22015996
-
A new genomic resource dedicated to wood formation in Eucalyptus.BMC Plant Biol. 2009 Mar 27;9:36. doi: 10.1186/1471-2229-9-36. BMC Plant Biol. 2009. PMID: 19327132 Free PMC article.
-
The ornamental variety, Japanese striped corn, contains high anthocyanin levels and PAL specific activity: establishing the potential for development of an oral therapeutic.Plant Cell Rep. 2009 Mar;28(3):503-15. doi: 10.1007/s00299-008-0650-6. Epub 2008 Dec 10. Plant Cell Rep. 2009. PMID: 19082600
-
A genomewide analysis of the cinnamyl alcohol dehydrogenase family in sorghum [Sorghum bicolor (L.) Moench] identifies SbCAD2 as the brown midrib6 gene.Genetics. 2009 Feb;181(2):783-95. doi: 10.1534/genetics.108.098996. Epub 2008 Dec 15. Genetics. 2009. PMID: 19087955 Free PMC article.
References
-
- Abdulrazzak N, Pollet B, Ehlting J, Larsen K, Asnaghi C, Ronseau S, Proux C, Erhardt M, Seltzer V, Renou JP, et al (2006) A coumaroyl-ester-3-hydroxylase insertion mutant reveals the existence of nonredundant meta-hydroxylation pathways and essential roles for phenolic precursors in cell expansion and plant growth. Plant Physiol 140 30–48 - PMC - PubMed
-
- Andjelkovic V, Thompson R (2006) Changes in gene expression in maize kernel in response to water and salt stress. Plant Cell Rep 25 71–79 - PubMed
-
- Appenzeller L, Doblin M, Barreiro R, Wang H, Niu X, Kollipara K (2004) Cellulose synthesis in maize: isolation and expression analysis of the cellulose synthase (CesA) gene family. Cellulose 11 287–299
-
- Arpat AB, Waugh M, Sullivan JP, Gonzales M, Frisch D, Main D, Wood T, Leslie A, Wing RA, Wilkins TA (2004) Functional genomics of cell elongation in developing cotton fibers. Plant Mol Biol 54 911–929 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
