

A chromatin-mediated mechanism for specification of conditional transcription factor targets
- PMID: 17099712
- PMCID: PMC2756100
- DOI: 10.1038/ng1917
A chromatin-mediated mechanism for specification of conditional transcription factor targets
Abstract
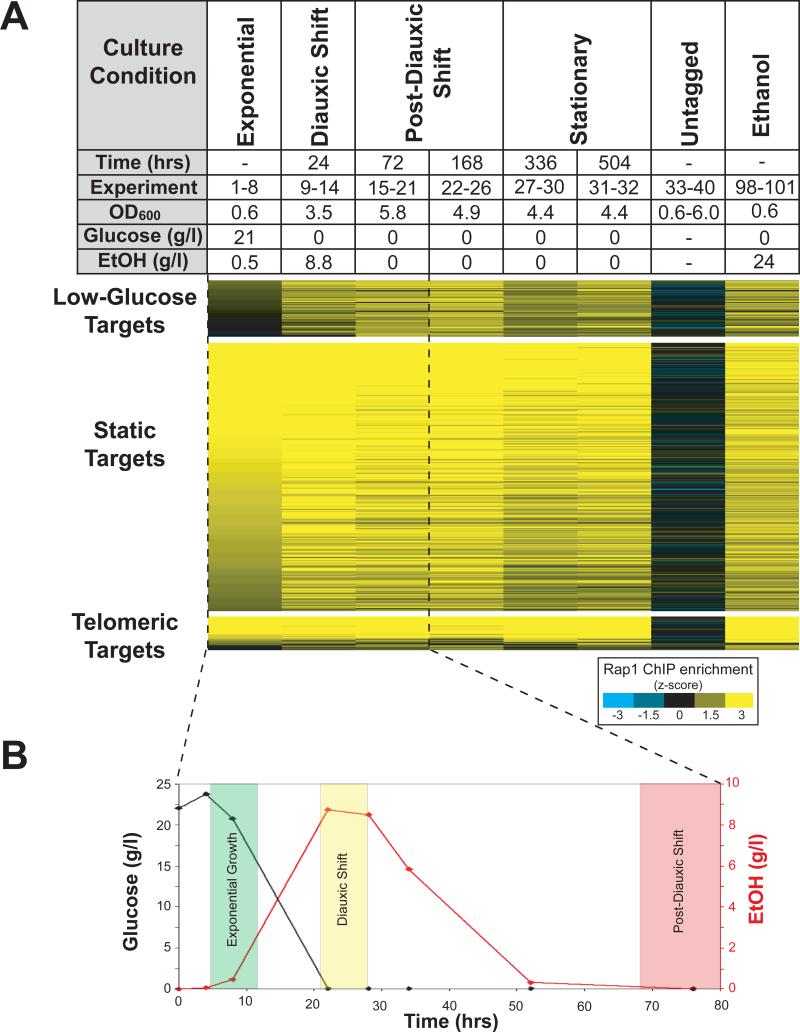
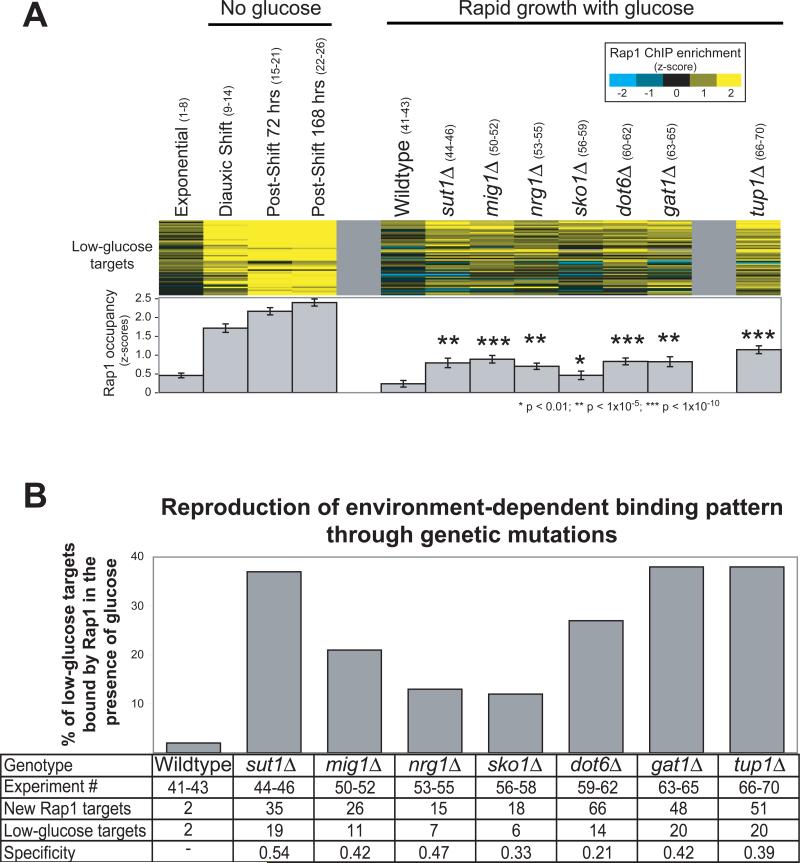
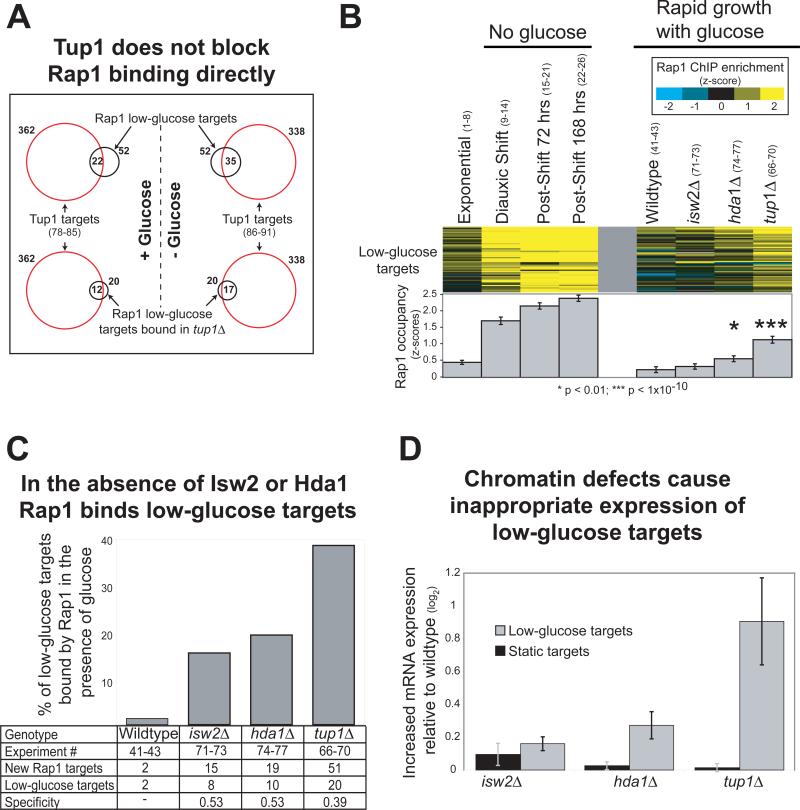
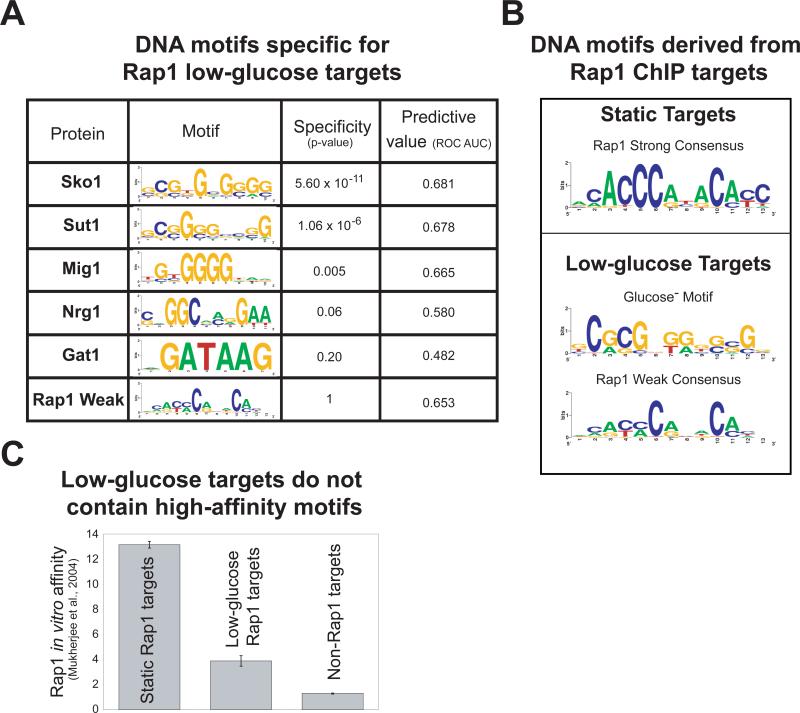
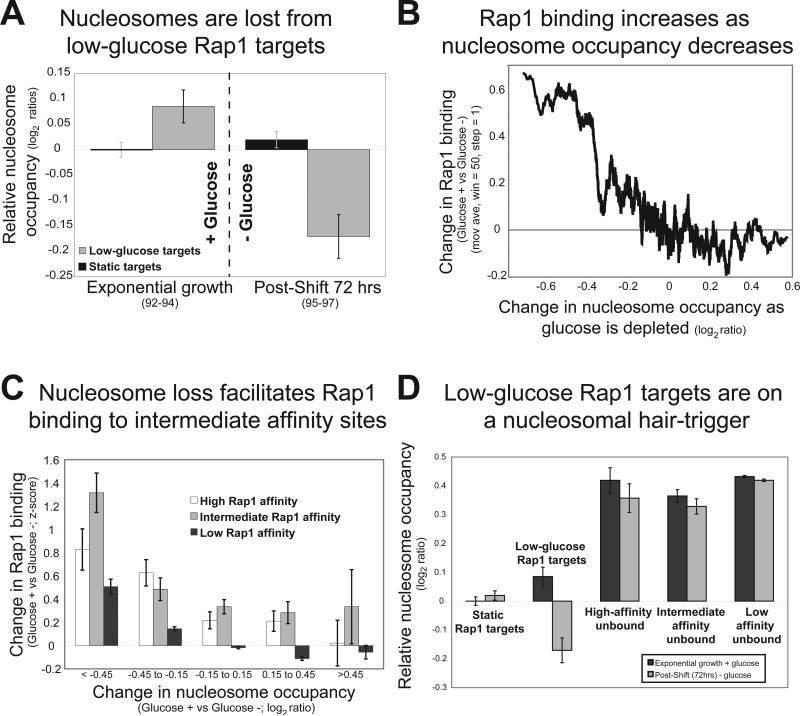
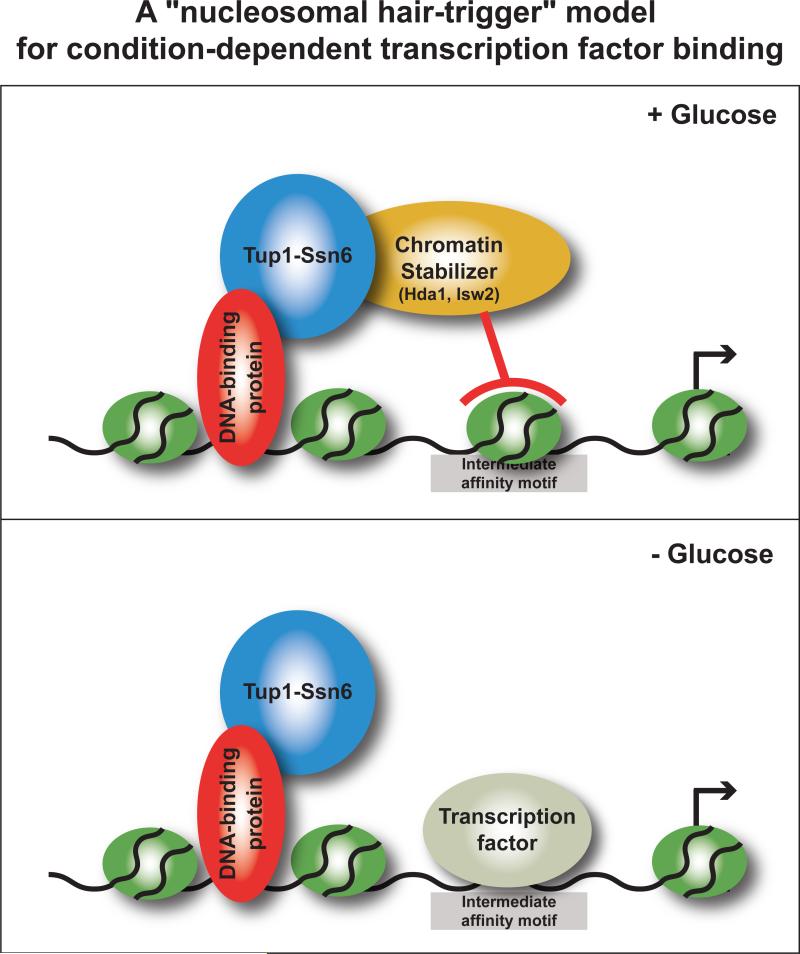
Organisms respond to changes in their environment, and many such responses are initiated at the level of gene transcription. Here, we provide evidence for a previously undiscovered mechanism for directing transcriptional regulators to new binding targets in response to an environmental change. We show that repressor-activator protein 1 (Rap1), a master regulator of yeast metabolism, binds to an expanded target set after glucose depletion despite decreasing protein levels and no evidence of posttranslational modification. Computational analysis predicts that proteins capable of recruiting the chromatin regulator Tup1 act to restrict the binding distribution of Rap1 in the presence of glucose. Deletion of the gene(s) encoding Tup1, recruiters of Tup1 or chromatin regulators recruited by Tup1 cause Rap1 to bind specifically and inappropriately to low-glucose targets. These data, combined with whole-genome measurements of nucleosome occupancy and Tup1 distribution, provide evidence for a mechanism of dynamic target specification that coordinates the genome-wide distribution of intermediate-affinity DNA sequence motifs with chromatin-mediated regulation of accessibility to those sites.
Figures
References
-
- Jacob F, Monod J. Genetic regulatory mechanisms in the synthesis of proteins. J Mol Biol. 1961;3:318–56. - PubMed
-
- O'Neill EM, Kaffman A, Jolly ER, O'Shea EK. Regulation of PHO4 nuclear localization by the PHO80-PHO85 cyclin-CDK complex. Science. 1996;271:209–12. - PubMed
-
- Gu W, Roeder RG. Activation of p53 sequence-specific DNA binding by acetylation of the p53 C-terminal domain. Cell. 1997;90:595–606. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
