

Molecular patterns of sex determination in the animal kingdom: a comparative study of the biology of reproduction
- PMID: 17101057
- PMCID: PMC1660543
- DOI: 10.1186/1477-7827-4-59
Molecular patterns of sex determination in the animal kingdom: a comparative study of the biology of reproduction
Abstract
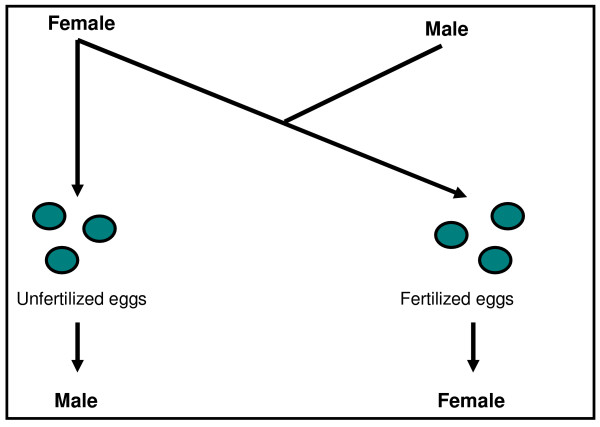
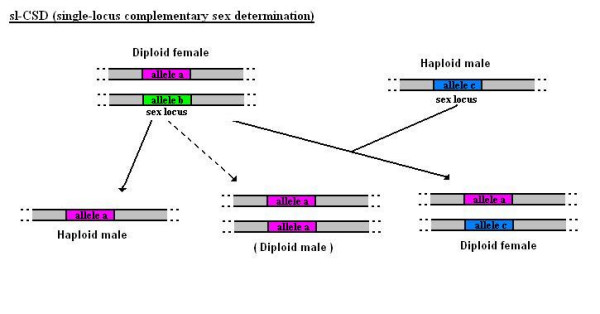
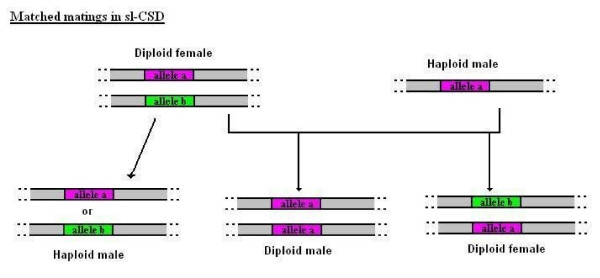
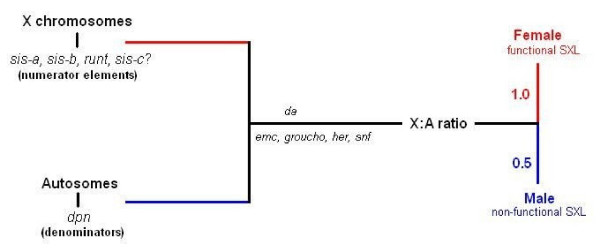
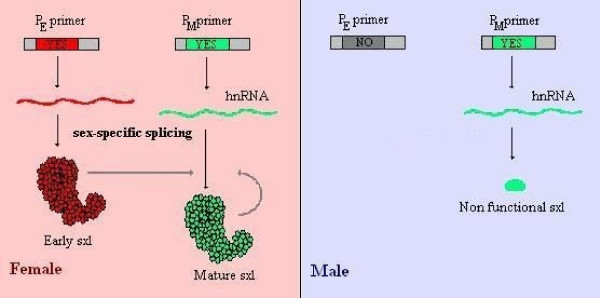
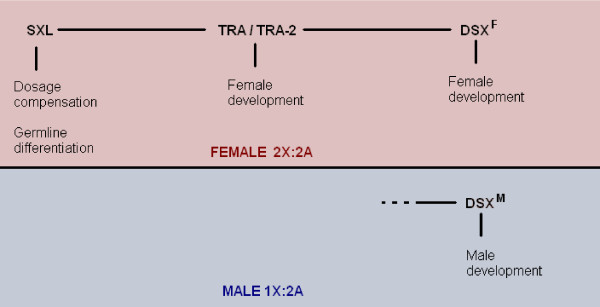
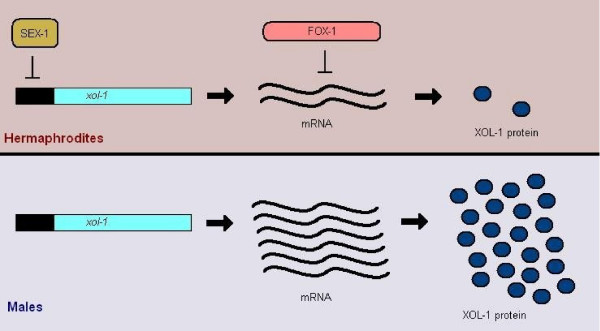
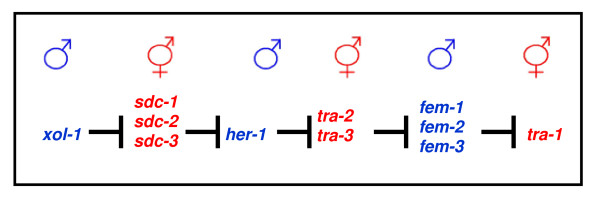
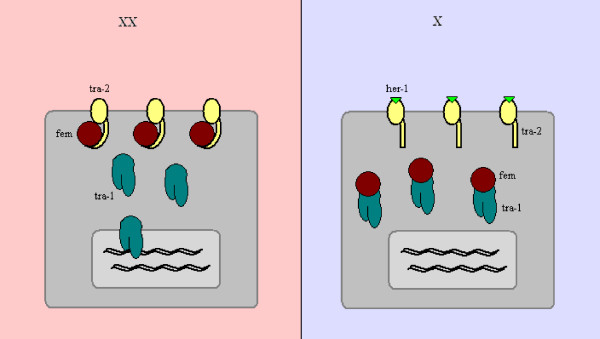
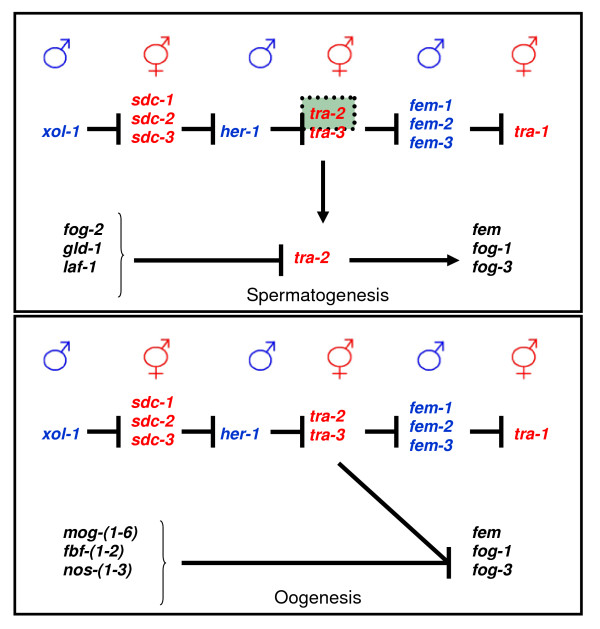
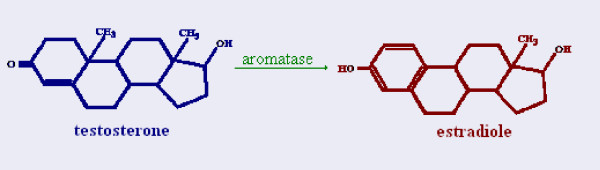
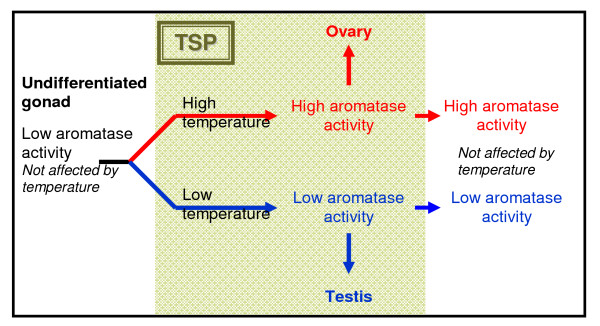
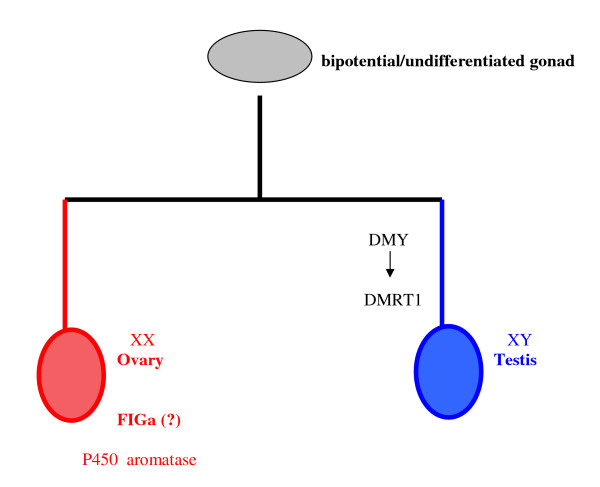
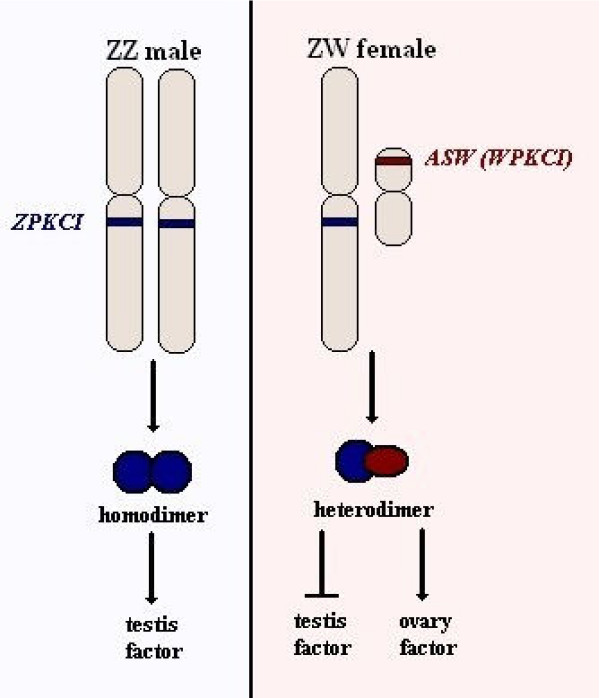
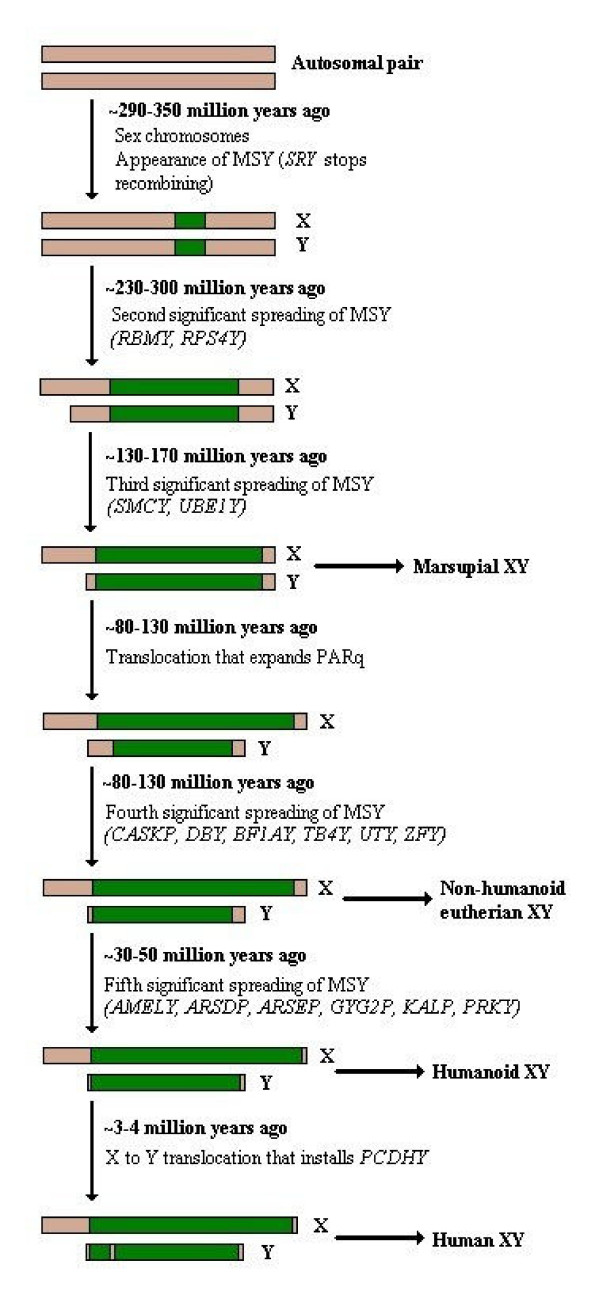
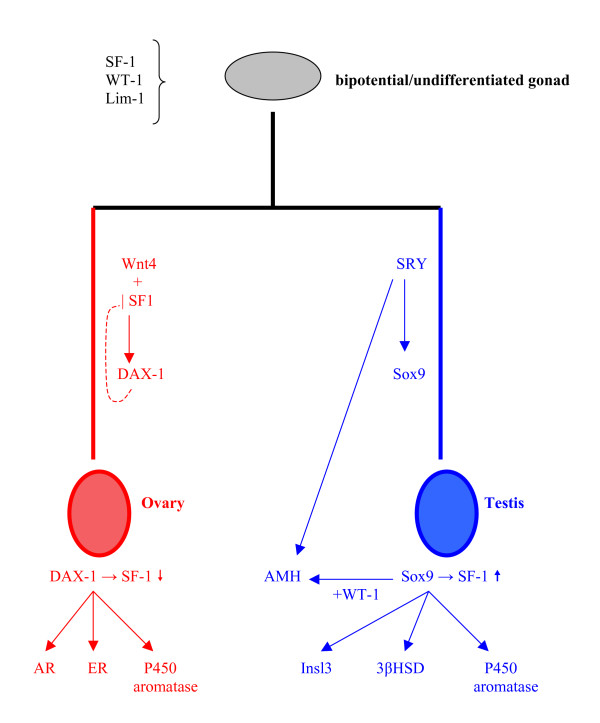
Determining sexual fate is an integral part of reproduction, used as a means to enrich the genome. A variety of such regulatory mechanisms have been described so far and some of the more extensively studied ones are being discussed. For the insect order of Hymenoptera, the choice lies between uniparental haploid males and biparental diploid females, originating from unfertilized and fertilized eggs accordingly. This mechanism is also known as single-locus complementary sex determination (slCSD). On the other hand, for Dipterans and Drosophila melanogaster, sex is determined by the ratio of X chromosomes to autosomes and the sex switching gene, sxl. Another model organism whose sex depends on the X:A ratio, Caenorhabditis elegans, has furthermore to provide for the brief period of spermatogenesis in hermaphrodites (XX) without the benefit of the "male" genes of the sex determination pathway. Many reptiles have no discernible sex determining genes. Their sexual fate is determined by the temperature of the environment during the thermosensitive period (TSP) of incubation, which regulates aromatase activity. Variable patterns of sex determination apply in fish and amphibians. In birds, while sex chromosomes do exist, females are the heterogametic (ZW) and males the homogametic sex (ZZ). However, we have yet to decipher which of the two (Z or W) is responsible for the choice between males and females. In mammals, sex determination is based on the presence of two identical (XX) or distinct (XY) gonosomes. This is believed to be the result of a lengthy evolutionary process, emerging from a common ancestral autosomal pair. Indeed, X and Y present different levels of homology in various mammals, supporting the argument of a gradual structural differentiation starting around the SRY region. The latter initiates a gene cascade that results in the formation of a male. Regulation of sex steroid production is also a major result of these genetic interactions. Similar observations have been described not only in mammals, but also in other vertebrates, emphasizing the need for further study of both normal hormonal regulators of sexual phenotype and patterns of epigenetic/environmental disruption.
Figures
References
-
- Manolakou P, Angelopoulou R, Lavranos G. Sex Determinants in the Genome: Lessons from the Animal Kingdom. Coll Antropol. 2006;30:649–52. - PubMed
-
- Howard J. Mitochondrial Eve, Y chromosome Adam, testosterone and human evolution. Riv Biol. 2002;95:319–325. - PubMed
-
- Lavranos G, Angelopoulou R, Manolakou P, Balla M. Hormonal and Meta-Hormonal Determinants of Sexual Dimorphism. Coll Antropol. 2006;30:659–63. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
