

Environmental mimics and the Lvh type IVA secretion system contribute to virulence-related phenotypes of Legionella pneumophila
- PMID: 17101653
- PMCID: PMC1828514
- DOI: 10.1128/IAI.00956-06
Environmental mimics and the Lvh type IVA secretion system contribute to virulence-related phenotypes of Legionella pneumophila
Abstract
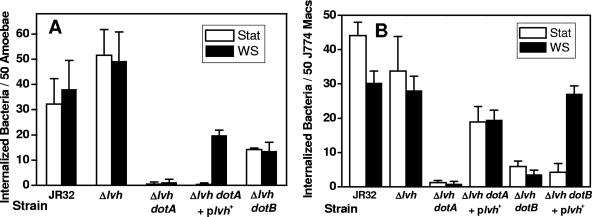
Legionella pneumophila, the causative organism of Legionnaires' disease, is a fresh-water bacterium and intracellular parasite of amoebae. This study examined the effects of incubation in water and amoeba encystment on L. pneumophila strain JR32 and null mutants in dot/icm genes encoding a type IVB secretion system required for entry, delayed acidification of L. pneumophila-containing phagosomes, and intracellular multiplication when stationary-phase bacteria infect amoebae and macrophages. Following incubation of stationary-phase cultures in water, mutants in dotA and dotB, essential for function of the type IVB secretion system, exhibited entry and delay of phagosome acidification comparable to that of strain JR32. Following encystment in Acanthamoeba castellanii and reversion of cysts to amoeba trophozoites, dotA and dotB mutants exhibited intracellular multiplication in amoebae. The L. pneumophila Lvh locus, encoding a type IVA secretion system homologous to that in Agrobacterium tumefaciens, was required for restoration of entry and intracellular multiplication in dot/icm mutants following incubation in water and amoeba encystment and was required for delay of phagosome acidification in strain JR32. These data support a model in which the Dot/Icm type IVB secretion system is conditionally rather than absolutely required for L. pneumophila virulence-related phenotypes. The data suggest that the Lvh type IVA secretion system, previously thought to be dispensable, is involved in virulence-related phenotypes under conditions mimicking the spread of Legionnaires' disease from environmental niches. Since environmental amoebae are implicated as reservoirs for an increasing number of environmental pathogens and for drug-resistant bacteria, the environmental mimics developed here may be useful in virulence studies of other pathogens.
Figures





References
-
- Abd, H., A. Weintraub, and G. Sandstrom. 2005. Intracellular survival and replication of Vibrio cholerae O139 in aquatic free-living amoebae. Environ. Microbiol. 7:1003-1008. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
