

Interleukin-17 is a negative regulator of established allergic asthma
- PMID: 17101734
- PMCID: PMC2118159
- DOI: 10.1084/jem.20061401
Interleukin-17 is a negative regulator of established allergic asthma
Abstract
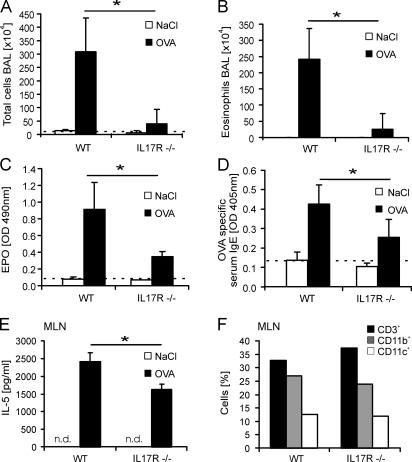
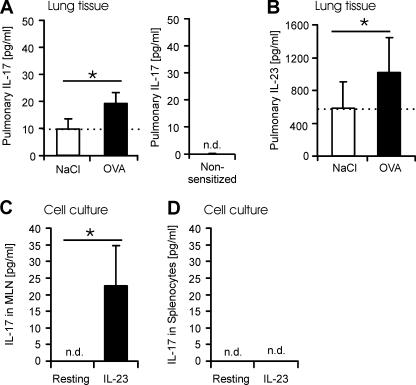
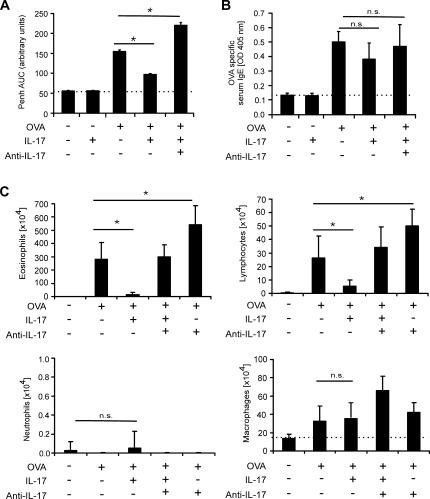
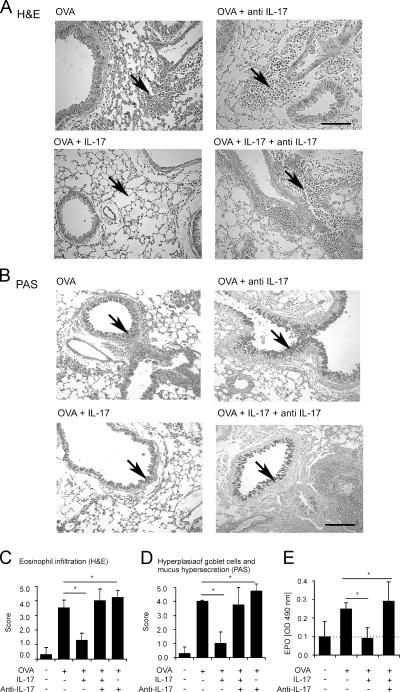
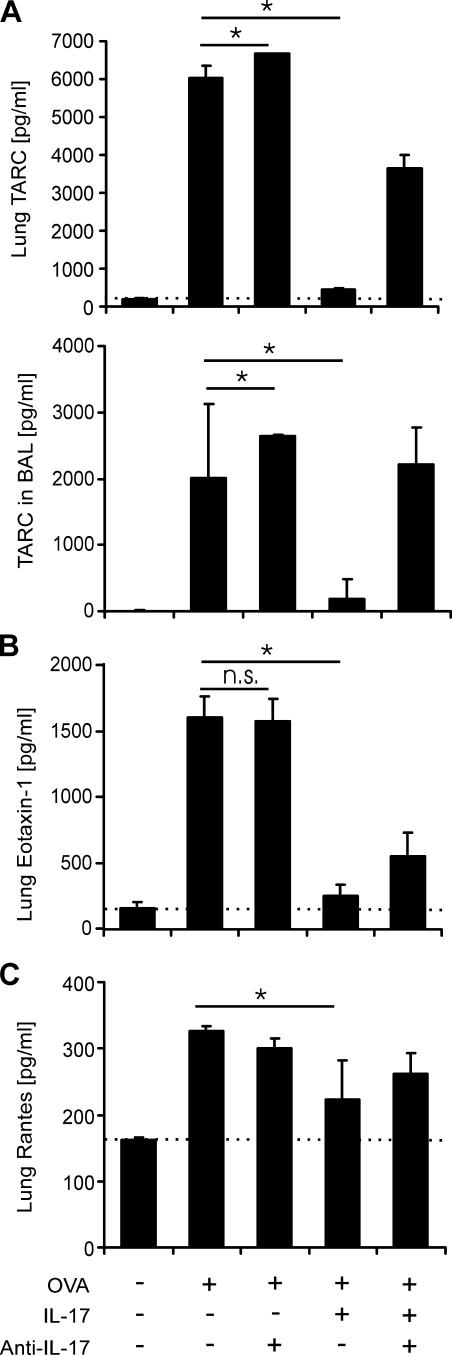
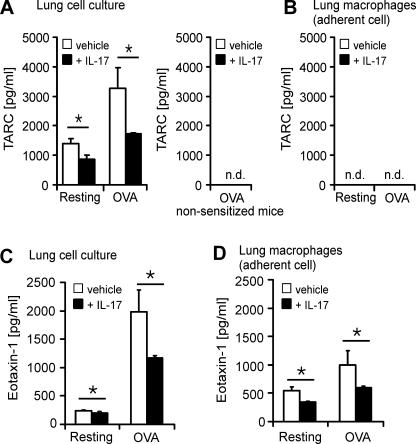
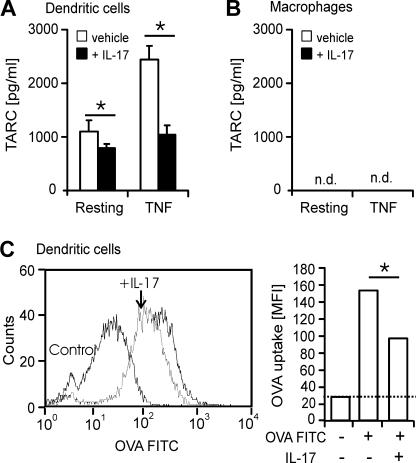
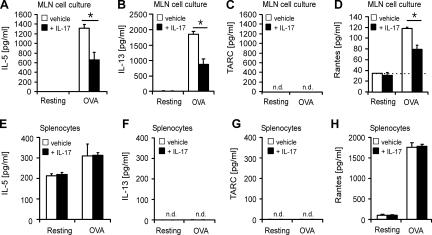
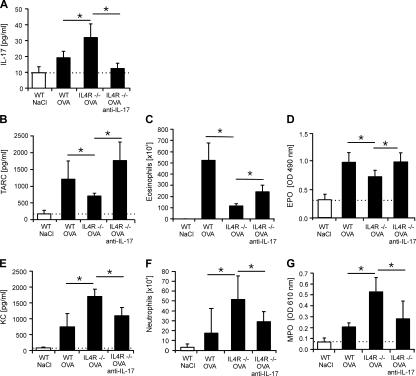
T helper (Th)17 cells producing interleukin (IL)-17 play a role in autoimmune and allergic inflammation. Here, we show that IL-23 induces IL-17 in the lung and IL-17 is required during antigen sensitization to develop allergic asthma, as shown in IL-17R-deficient mice. Since IL-17 expression increased further upon antigen challenge, we addressed its function in the effector phase. Most strikingly, neutralization of IL-17 augmented the allergic response in sensitized mice. Conversely, exogenous IL-17 reduced pulmonary eosinophil recruitment and bronchial hyperreactivity, demonstrating a novel regulatory role of IL-17. Mechanistically, IL-17 down modulated eosinophil-chemokine eotaxin (CCL11) and thymus- and activation-regulated chemokine/CCL17 (TARC) in lungs in vivo and ex vivo upon antigen restimulation. In vitro, IL-17 reduced TARC production in dendritic cells (DCs)-the major source of TARC-and antigen uptake by DCs and IL-5 and IL-13 production in regional lymph nodes. Furthermore, IL-17 is regulated in an IL-4-dependent manner since mice deficient for IL-4Ralpha signaling showed a marked increase in IL-17 concentration with inhibited eosinophil recruitment. Therefore, endogenous IL-17 is controlled by IL-4 and has a dual role. Although it is essential during antigen sensitization to establish allergic asthma, in sensitized mice IL-17 attenuates the allergic response by inhibiting DCs and chemokine synthesis.
Figures
References
-
- Wills-Karp, M. 1999. Immunologic basis of antigen-induced airway hyperresponsiveness. Annu. Rev. Immunol. 17:255–281. - PubMed
-
- D'Ambrosio, D., A. Iellem, R. Bonecchi, D. Mazzeo, S. Sozzani, A. Mantovani, and F. Sinigaglia. 1998. Selective up-regulation of chemokine receptors CCR4 and CCR8 upon activation of polarized human type 2 Th cells. J. Immunol. 161:5111–5115. - PubMed
-
- Herrick, C.A., and K. Bottomly. 2003. To respond or not to respond: T cells in allergic asthma. Nat. Rev. Immunol. 3:405–412. - PubMed
-
- Fossiez, F., J. Banchereau, R. Murray, C. Van Kooten, P. Garrone, and S. Lebecque. 1998. Interleukin-17. Int. Rev. Immunol. 16:541–551. - PubMed
-
- Yao, Z., S.L. Painter, W.C. Fanslow, D. Ulrich, B.M. Macduff, M.K. Spriggs, and R.J. Armitage. 1995. Human IL-17: a novel cytokine derived from T cells. J. Immunol. 155:5483–5486. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
