

Spatial and nonspatial escape strategies in the Barnes maze
- PMID: 17101874
- PMCID: PMC1783636
- DOI: 10.1101/lm.334306
Spatial and nonspatial escape strategies in the Barnes maze
Abstract
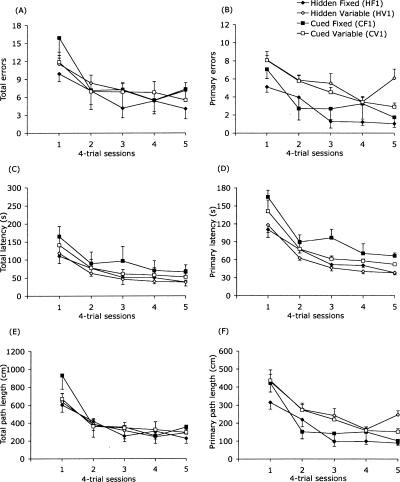
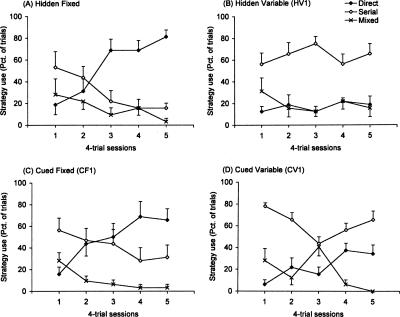
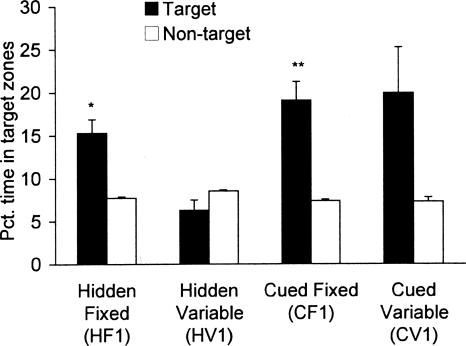
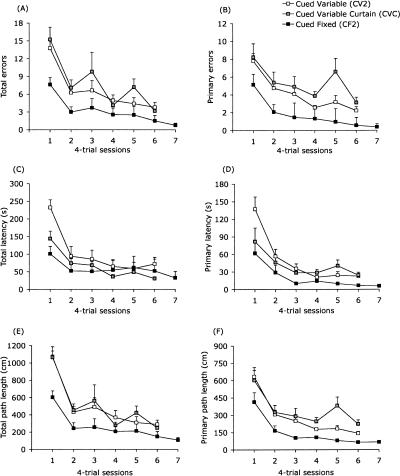
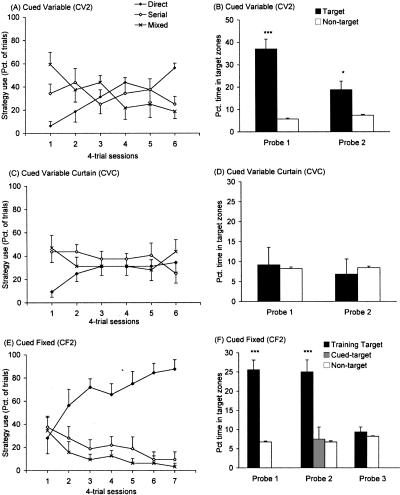
The Barnes maze is a spatial memory task that requires subjects to learn the position of a hole that can be used to escape the brightly lit, open surface of the maze. Two experiments assessed the relative importance of spatial (extra-maze) versus proximal visible cues in solving the maze. In Experiment 1, four groups of mice were trained either with or without a discrete visible cue marking the location of the escape hole, which was either in a fixed or variable location across trials. In Experiment 2, all mice were trained with the discrete visible cue marking the target hole location. Two groups were identical to the cued-target groups from Experiment 1, with either fixed or variable escape locations. For these mice, the discrete cue either was the sole predictor of the target location or was perfectly confounded with the spatial extra-maze cues. The third group also used a cued variable target, but a curtain was drawn around the maze to prevent the use of spatial cues to guide navigation. Probe trials with all escape holes blocked were conducted to dissociate the use of spatial and discrete proximal cues. We conclude that the Barnes maze can be solved efficiently using spatial, visual cue, or serial-search strategies. However, mice showed a strong preference for using the distal room cues, even when a discrete visible cue clearly marked the escape location. Importantly, these data show that the cued-target control version of the Barnes maze as typically conducted does not dissociate spatial from nonspatial abilities.
Figures
References
-
- Barnes C.A. Memory deficits associated with senescence: A neurophysiological and behavioral study in the rat. J. Comp. Physiol. Psychol. 1979;93:74–104. - PubMed
-
- Bernardo A., McCord M., Troen A.M., Allison J.D., McDonald M.P. Impaired spatial memory in APP-overexpressing mice on a homocysteinemia-inducing diet. Neurobiol. Aging. 2006 (in press) - PubMed
-
- Biegler R., Morris R.G. Landmark stability is a prerequisite for spatial but not discrimination learning. Nature. 1993;361:631–633. - PubMed
-
- Biegler R., Morris R.G. Landmark stability: Further studies pointing to a role in spatial learning. Q. J. Exp. Psychol. B. 1996;49:307–345. - PubMed
-
- Biegler R., Morris R.G. Blocking in the spatial domain with arrays of discrete landmarks. J. Exp. Psychol. Anim. Behav. Process. 1999;25:334–351. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources