

Na channel inactivation from open and closed states
- PMID: 17101981
- PMCID: PMC1693860
- DOI: 10.1073/pnas.0607603103
Na channel inactivation from open and closed states
Abstract
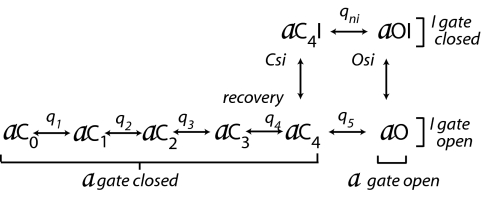
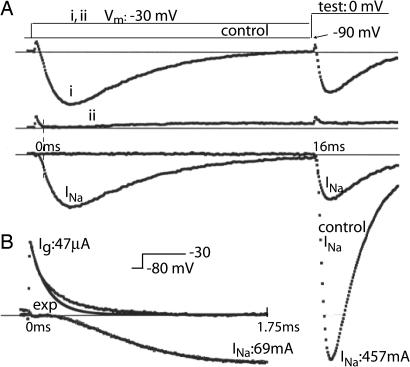
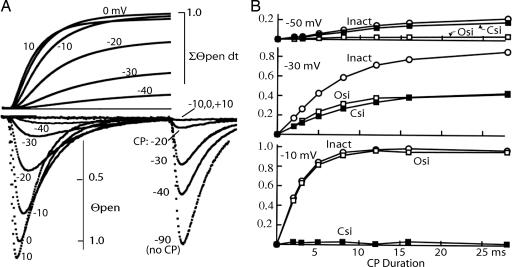
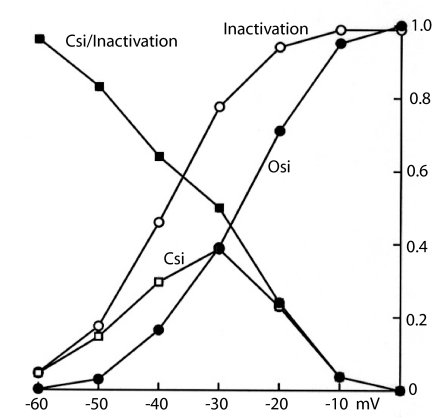
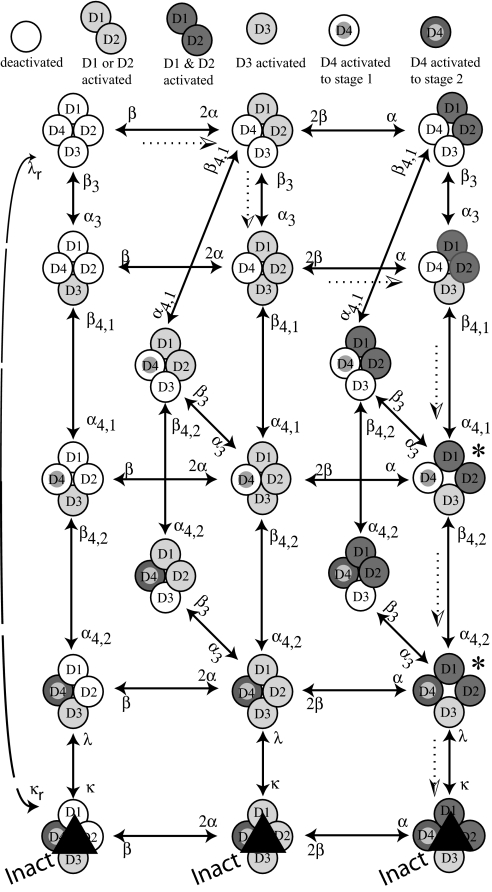
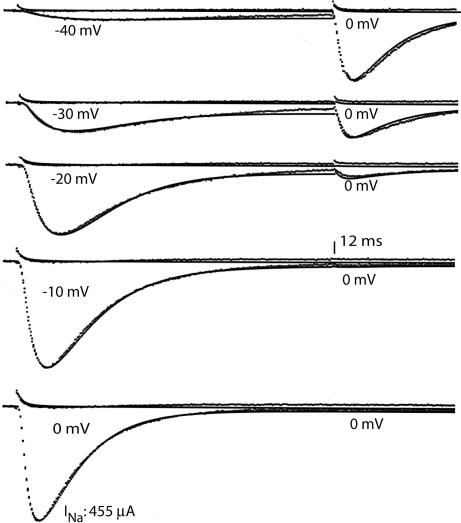
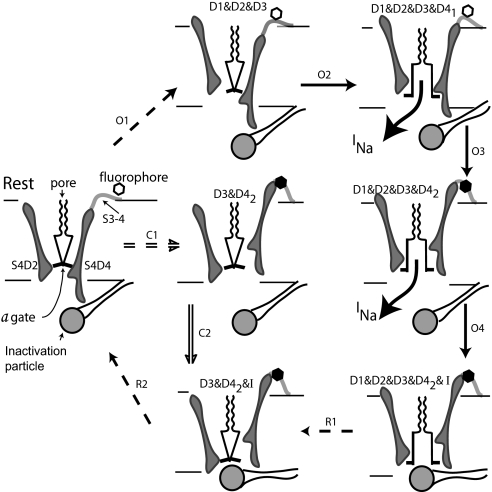
A sodium channel is composed of four similar domains, each containing a highly charged S4 helix that is driven outward (activates) in response to a depolarization. Functionally, the channel has two gates, called activation gate (a gate) and inactivation gate (I gate), both of which must be open for conduction to occur. The cytoplasmically located a gate opens after a depolarization has activated the S4s of (probably) all four domains. The I gate consists of a cytoplasmically located inactivation "particle" and a receptor for it in the channel. The receptor becomes available after some degree of S4 activation, and the particle diffuses in to inactivate the channel. The I gate usually closes when the a gate is open [open-state inactivation (Osi)] but also can close before the channel reaches the conducting state. This "closed-state inactivation" (Csi) is studied quantitatively in this paper to determine the degree of S4 activation required for (i) opening the a gate, and (ii) permitting the I gate to close. Csi is most prominent for small depolarizations, during which occupancy of the partially activated closed states is prolonged. Large depolarizations drive the S4s outward quickly, minimizing the duration of closed-state occupancy and making Csi small and Osi large. Based on these data and evidence in the literature, it is concluded that opening the a gate requires S4 activation in domains 1-3, with partial activation of the S4 of domain 4. Csi requires only S4 activation of domains 3 and 4, which does not open the a gate.
Conflict of interest statement
The author declares no conflict of interest.
Figures
References
-
- Aldrich RW, Corey DP, Stevens CF. Nature. 1983;306:436–441. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
