

Micromolar intracellular hydrogen peroxide disrupts metabolism by damaging iron-sulfur enzymes
- PMID: 17102132
- PMCID: PMC5136138
- DOI: 10.1074/jbc.M607646200
Micromolar intracellular hydrogen peroxide disrupts metabolism by damaging iron-sulfur enzymes
Abstract
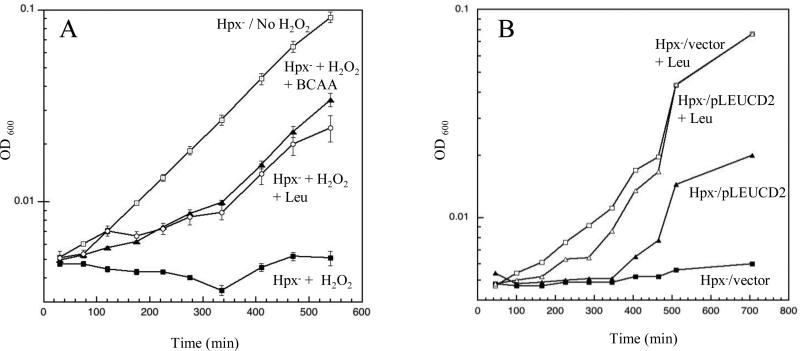
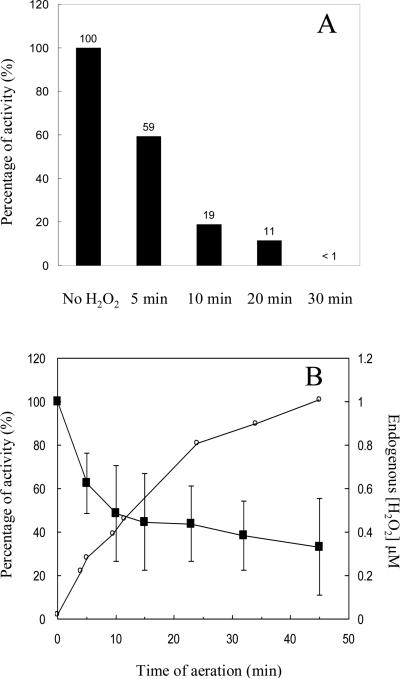
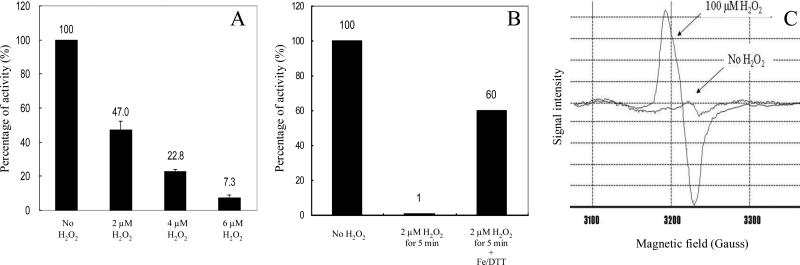
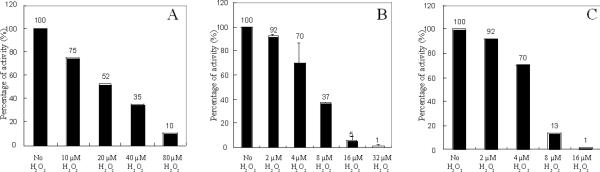
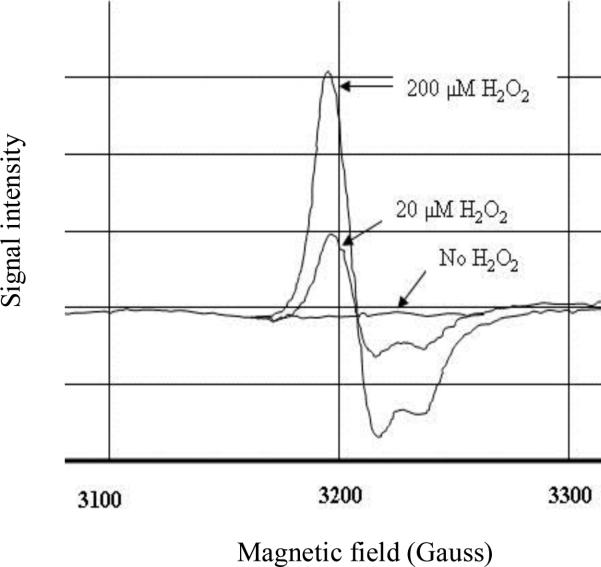
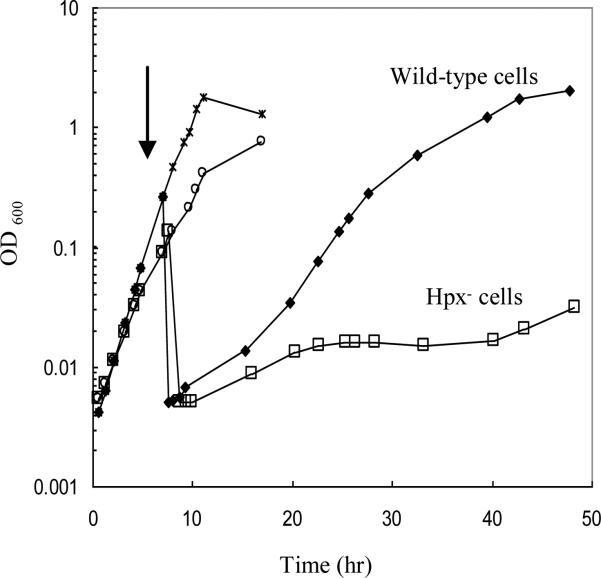
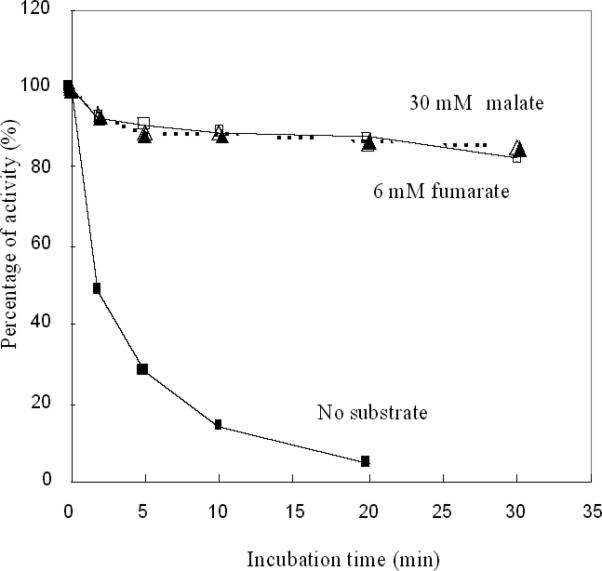
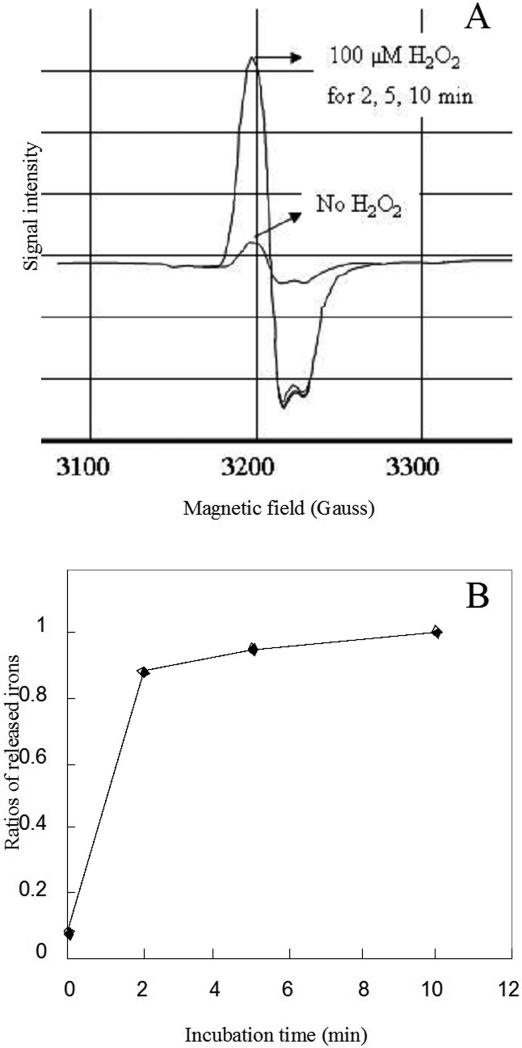
An Escherichia coli strain that cannot scavenge hydrogen peroxide has been used to identify the cell processes that are most sensitive to this oxidant. Low micromolar concentrations of H2O2 completely blocked the biosynthesis of leucine. The defect was tracked to the inactivation of isopropylmalate isomerase. This enzyme belongs to a family of [4Fe-4S] dehydratases that are notoriously sensitive to univalent oxidation, and experiments confirmed that other members were also inactivated. In vitro and in vivo analyses showed that H2O2 directly oxidized their solvent-exposed clusters in a Fenton-like reaction. The oxidized cluster then degraded to a catalytically inactive [3Fe-4S] form. Experiments indicated that H2O2 accepted two consecutive electrons during the oxidation event. As a consequence, hydroxyl radicals were not released; the polypeptide was undamaged; and the enzyme was competent for reactivation by repair processes. Strikingly, in scavenger-deficient mutants, the H2O2 that was generated as an adventitious by-product of metabolism (<1 microm) was sufficient to damage these [4Fe-4S] enzymes. This result demonstrates that aerobic organisms must synthesize H2O2 scavengers to avoid poisoning their own pathways. The extreme vulnerability of these enzymes may explain why many organisms, including mammals, deploy H2O2 to suppress microbial growth.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
