

A novel computational model of the circadian clock in Arabidopsis that incorporates PRR7 and PRR9
- PMID: 17102803
- PMCID: PMC1682023
- DOI: 10.1038/msb4100101
A novel computational model of the circadian clock in Arabidopsis that incorporates PRR7 and PRR9
Abstract
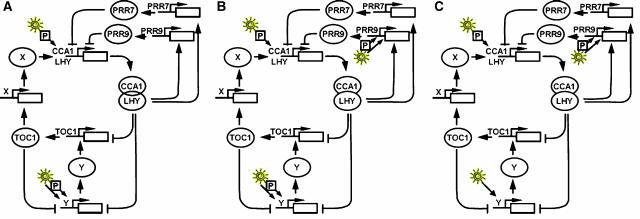
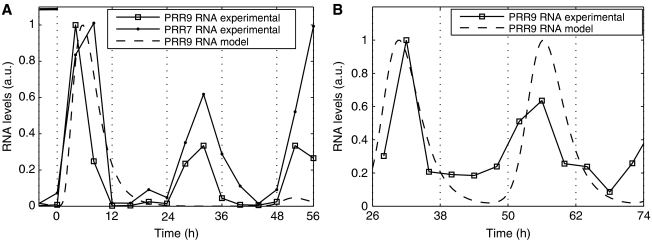
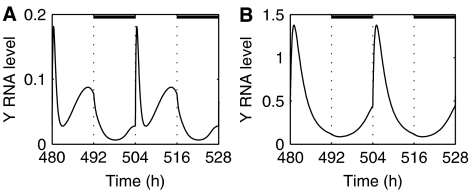
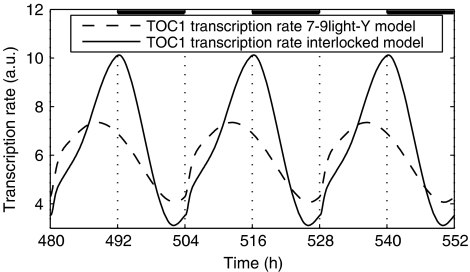
In plants, as in animals, the core mechanism to retain rhythmic gene expression relies on the interaction of multiple feedback loops. In recent years, molecular genetic techniques have revealed a complex network of clock components in Arabidopsis. To gain insight into the dynamics of these interactions, new components need to be integrated into the mathematical model of the plant clock. Our approach accelerates the iterative process of model identification, to incorporate new components, and to systematically test different proposed structural hypotheses. Recent studies indicate that the pseudo-response regulators PRR7 and PRR9 play a key role in the core clock of Arabidopsis. We incorporate PRR7 and PRR9 into an existing model involving the transcription factors TIMING OF CAB (TOC1), LATE ELONGATED HYPOCOTYL (LHY) and CIRCADIAN CLOCK ASSOCIATED (CCA1). We propose candidate models based on experimental hypotheses and identify the computational models with the application of an optimization routine. Validation is accomplished through systematic analysis of various mutant phenotypes. We introduce and apply sensitivity analysis as a novel tool for analyzing and distinguishing the characteristics of proposed architectures, which also allows for further validation of the hypothesized structures.
Figures

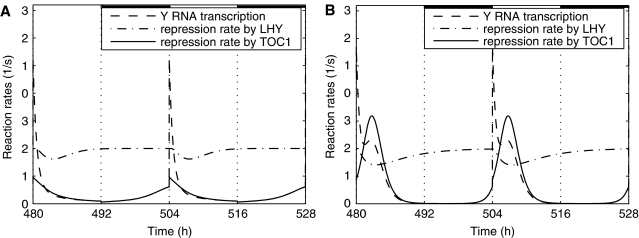
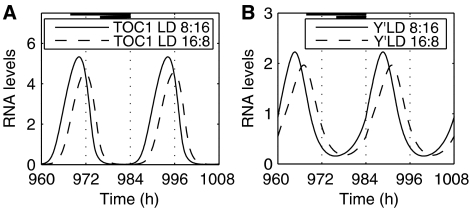
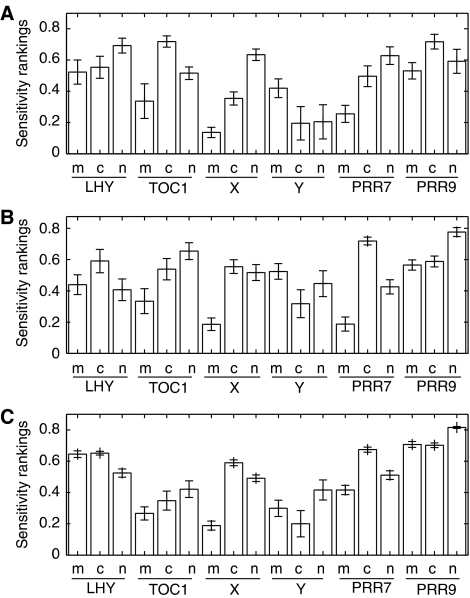
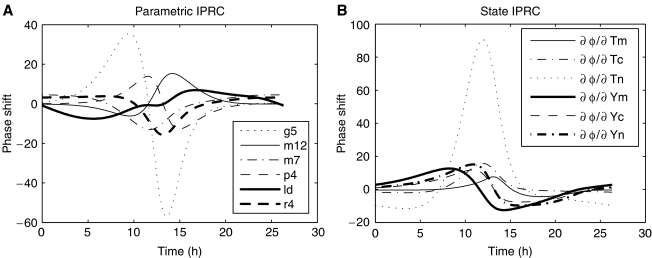
Comment in
-
Systems biology flowering in the plant clock field.Mol Syst Biol. 2006;2:60. doi: 10.1038/msb4100105. Epub 2006 Nov 14. Mol Syst Biol. 2006. PMID: 17102805 Free PMC article. No abstract available.
References
-
- Alabadi D, Oyama T, Yanovsky MJ, Harmon FG, Mas P, Kay SA (2001) Reciprocal regulation between TOC1 and LHY/CCA1 within the Arabidopsis circadian clock. Science 293: 880–883 - PubMed
-
- Alabadi D, Yanovsky MJ, Mas P, Harmer SL, Kay SA (2002) Critical role for CCA1 and LHY in maintaining circadian rhythmicity in Arabidopsis. Curr Biol 12: 757–761 - PubMed
-
- Csete ME, Doyle JC (2002) Reverse engineering of biological complexity. Science 295: 1664–1669 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
