

Experimental validation of a predicted feedback loop in the multi-oscillator clock of Arabidopsis thaliana
- PMID: 17102804
- PMCID: PMC1682024
- DOI: 10.1038/msb4100102
Experimental validation of a predicted feedback loop in the multi-oscillator clock of Arabidopsis thaliana
Abstract
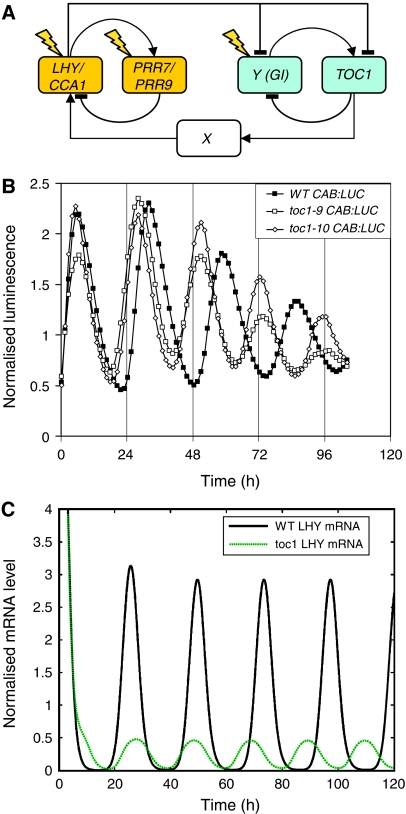
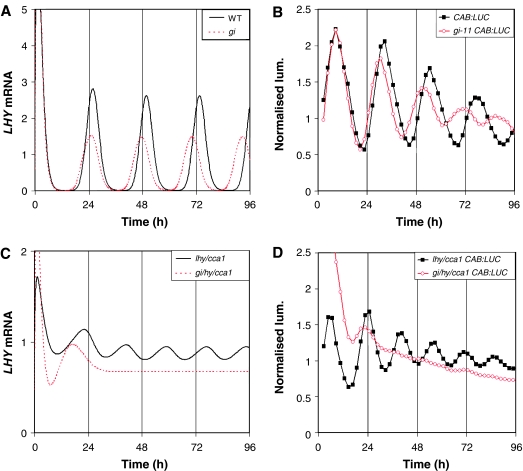
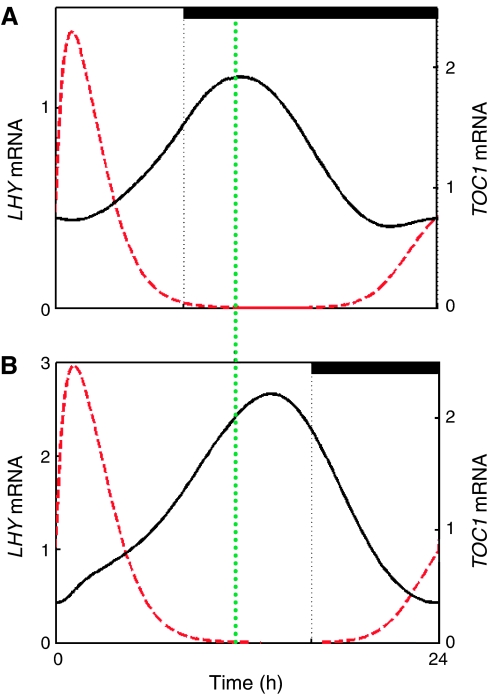
Our computational model of the circadian clock comprised the feedback loop between LATE ELONGATED HYPOCOTYL (LHY), CIRCADIAN CLOCK ASSOCIATED 1 (CCA1) and TIMING OF CAB EXPRESSION 1 (TOC1), and a predicted, interlocking feedback loop involving TOC1 and a hypothetical component Y. Experiments based on model predictions suggested GIGANTEA (GI) as a candidate for Y. We now extend the model to include a recently demonstrated feedback loop between the TOC1 homologues PSEUDO-RESPONSE REGULATOR 7 (PRR7), PRR9 and LHY and CCA1. This three-loop network explains the rhythmic phenotype of toc1 mutant alleles. Model predictions fit closely to new data on the gi;lhy;cca1 mutant, which confirm that GI is a major contributor to Y function. Analysis of the three-loop network suggests that the plant clock consists of morning and evening oscillators, coupled intracellularly, which may be analogous to coupled, morning and evening clock cells in Drosophila and the mouse.
Figures
Comment in
-
Systems biology flowering in the plant clock field.Mol Syst Biol. 2006;2:60. doi: 10.1038/msb4100105. Epub 2006 Nov 14. Mol Syst Biol. 2006. PMID: 17102805 Free PMC article. No abstract available.
References
-
- Alabadi D, Oyama T, Yanovsky MJ, Harmon FG, Mas P, Kay SA (2001) Reciprocal regulation between TOC1 and LHY/CCA1 within the Arabidopsis circadian clock. Science 293: 880–883 - PubMed
-
- Alabadi D, Yanovsky MJ, Mas P, Harmer SL, Kay SA (2002) Critical role for CCA1 and LHY in maintaining circadian rhythmicity in arabidopsis. Curr Biol 12: 757–761 - PubMed
-
- Duffield GE (2003) DNA microarray analyses of circadian timing: the genomic basis of biological time. J Neuroendocrinol 15: 991–1002 - PubMed
-
- Dunlap JC, Loros JJ, De Coursey PJ (2003) Chronobiology: biological timekeeping. Sunderland, MA, USA: Sinauer Associates
-
- Farre EM, Harmer SL, Harmon FG, Yanovsky MJ, Kay SA (2005) Overlapping and distinct roles of PRR7 and PRR9 in the Arabidopsis circadian clock. Curr Biol 15: 47–54 - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
