

No promoter left behind: global circadian gene expression in cyanobacteria
- PMID: 17107933
- PMCID: PMC3431602
- DOI: 10.1177/0748730406294418
No promoter left behind: global circadian gene expression in cyanobacteria
Abstract
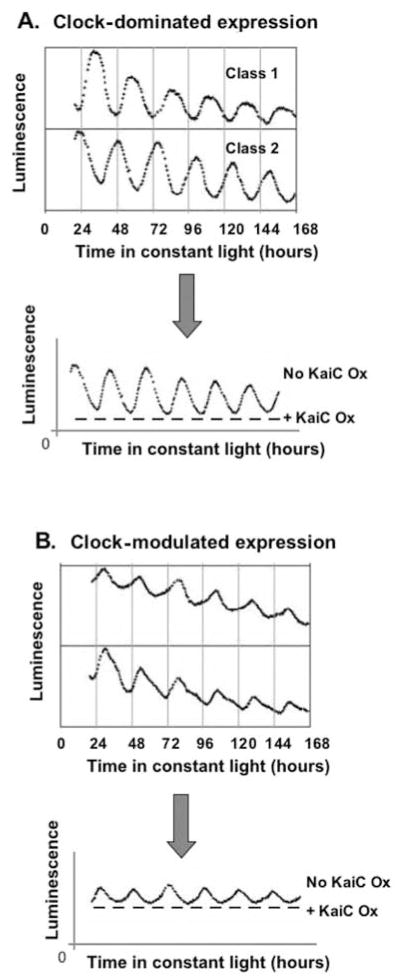
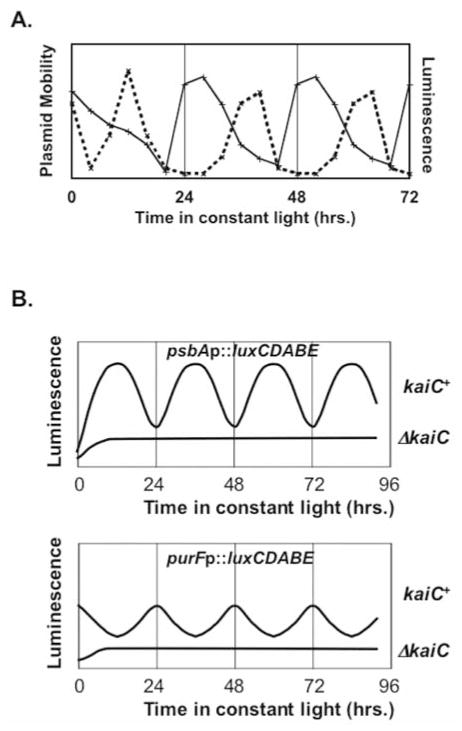
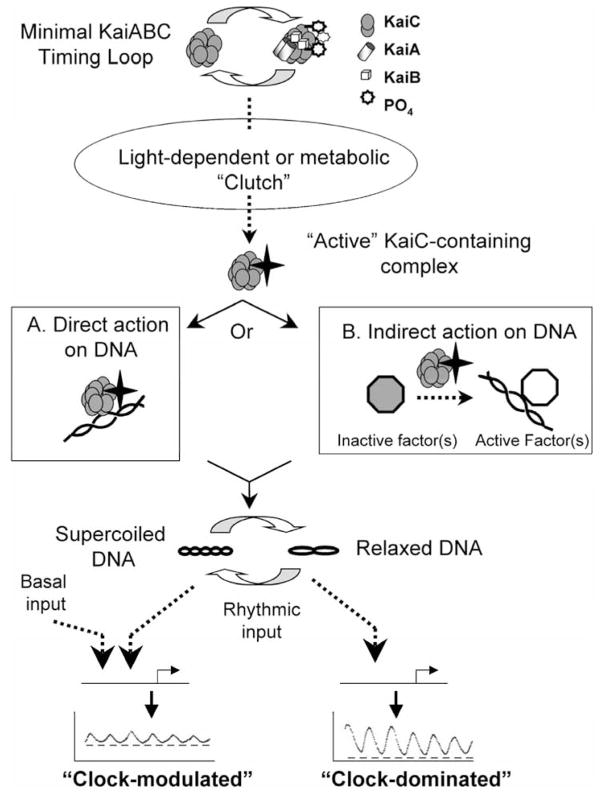
Prokaryotic cyanobacteria express robust circadian (daily) rhythms under the control of a clock system that appears to be similar to those of eukaryotes in many ways. On the other hand, the KaiABC-based core cyanobacterial clockwork is clearly different from the transcription-translation feedback loop model of eukaryotic clocks in that the cyanobacterial clock system regulates gene expression patterns globally, and specific clock gene promoters are not essential in mediating the circadian feedback loop. A novel model, the oscilloid model, proposes that the KaiABC oscillator ultimately mediates rhythmic changes in the status of the cyanobacterial chromosome, and these topological changes underlie the global rhythms of transcription. The authors suggest that this model represents one of several possible modes of regulating gene expression by circadian clocks, even those of eukaryotes.
Figures
References
-
- Akhtar RA, Reddy AB, Maywood ES, Clayton JD, King VM, Smith AG, Gant TW, Hastings MH, Kyriacou CP. Circadian cycling of the mouse liver transcriptome, as revealed by cDNA microarray, is driven by the suprachiasmatic nucleus. Curr Biol. 2002;12:540–550. - PubMed
-
- Chen YB, Dominic B, Zani S, Mellon MT, Zehr JP. Expression of photosynthesis genes in relation to nitrogen fixation in the diazotrophic filamentous nonheterocystous cyanobacterium Trichodesmium sp. IMS 101. Plant Mol Biol. 1999;41:89–104. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
