

The Old World and New World alphaviruses use different virus-specific proteins for induction of transcriptional shutoff
- PMID: 17108023
- PMCID: PMC1865960
- DOI: 10.1128/JVI.02073-06
The Old World and New World alphaviruses use different virus-specific proteins for induction of transcriptional shutoff
Abstract
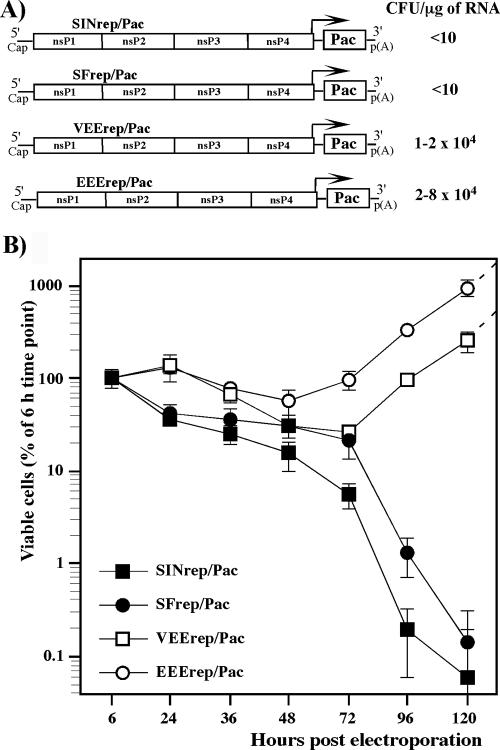
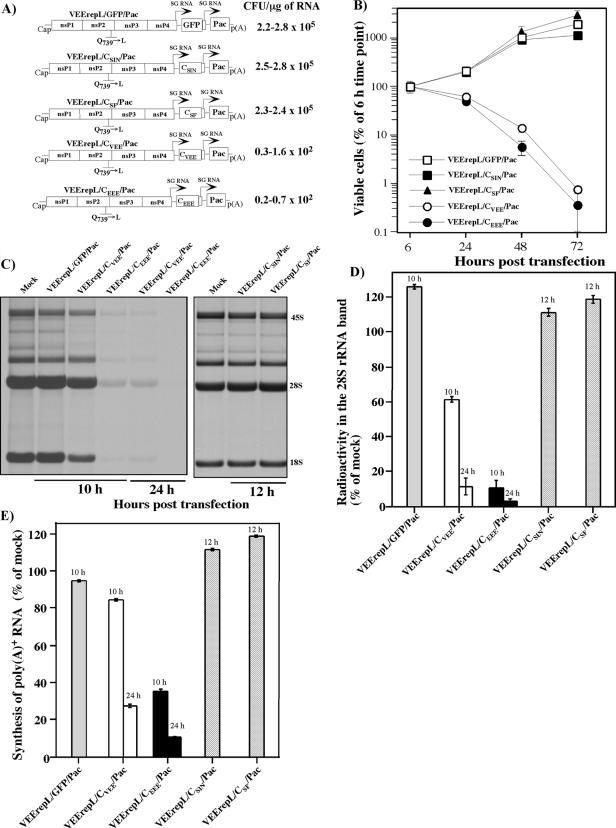
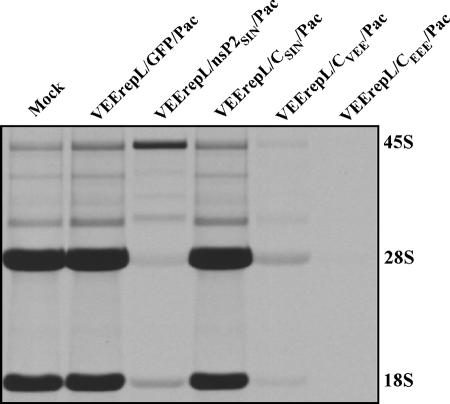
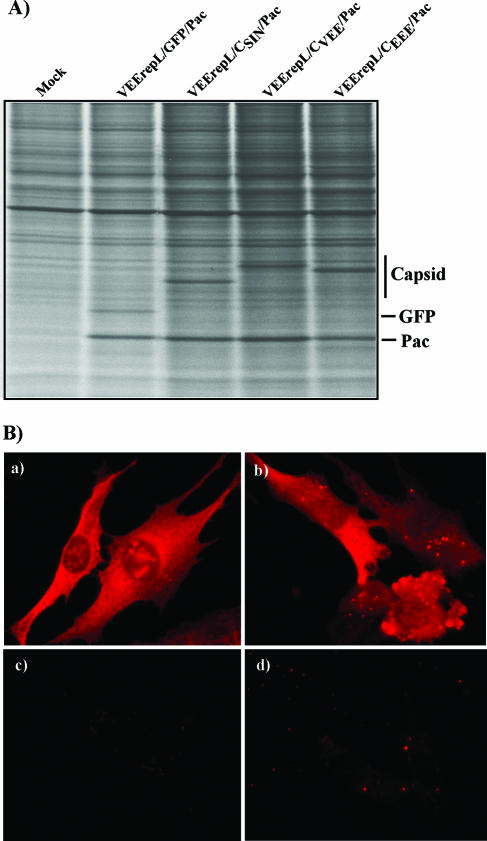
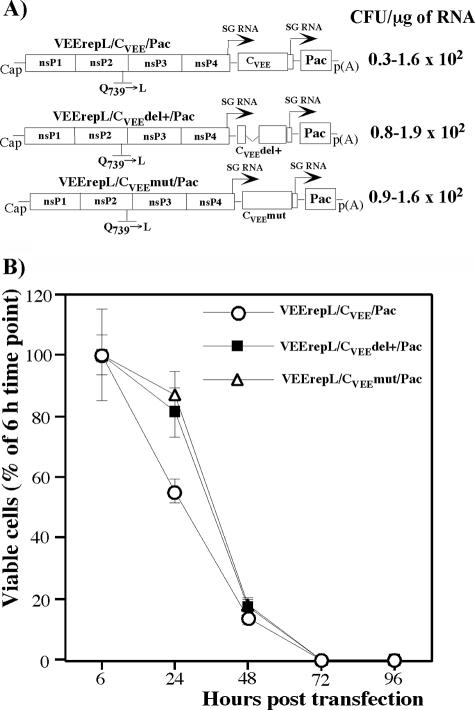
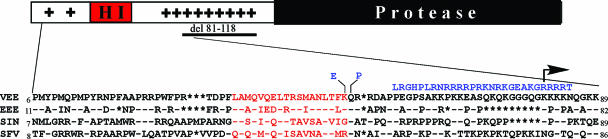
Alphaviruses are widely distributed throughout the world. During the last few thousand years, the New World viruses, including Venezuelan equine encephalitis virus (VEEV) and eastern equine encephalitis virus (EEEV), evolved separately from those of the Old World, i.e., Sindbis virus (SINV) and Semliki Forest virus (SFV). Nevertheless, the results of our study indicate that both groups have developed the same characteristic: their replication efficiently interferes with cellular transcription and the cell response to virus replication. Transcriptional shutoff caused by at least two of the Old World alphaviruses, SINV and SFV, which belong to different serological complexes, depends on nsP2, but not on the capsid protein, functioning. Our data suggest that the New World alphaviruses VEEV and EEEV developed an alternative mechanism of transcription inhibition that is mainly determined by their capsid protein, but not by the nsP2. The ability of the VEEV capsid to inhibit cellular transcription appears to be controlled by the amino-terminal fragment of the protein, but not by its protease activity or by the positively charged RNA-binding domain. These data provide new insights into alphavirus evolution and present a plausible explanation for the particular recombination events that led to the formation of western equine encephalitis virus (WEEV) from SINV- and EEEV-like ancestors. The recombination allowed WEEV to acquire capsid protein functioning in transcription inhibition from EEEV-like virus. Identification of the new functions in the New World alphavirus-derived capsids opens an opportunity for developing new, safer alphavirus-based gene expression systems and designing new types of attenuated vaccine strains of VEEV and EEEV.
Figures

References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
