

Immunoglobulin G signaling activates lysosome/phagosome docking
- PMID: 17110435
- PMCID: PMC1838734
- DOI: 10.1073/pnas.0609182103
Immunoglobulin G signaling activates lysosome/phagosome docking
Abstract
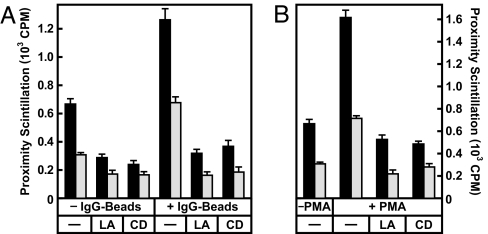
An important role of IgG antibodies in the defense against microbial infections is to promote the ingestion and killing of microbes by phagocytes. Here, we developed in vivo and in vitro approaches to ask whether opsonization of particles with IgG enhances intracellular targeting of lysosomes to phagosomes. To eliminate the effect of IgG on the ingestion process, cells were exposed to latex beads at 15-20 degrees C, which allows engulfment of both IgG-coated and uncoated beads but prevents the fusion of lysosomes with phagosomes. Upon shifting the temperature to 37 degrees C, phagosomes containing IgG beads matured significantly faster into phagolysosomes as judged by colocalization with lysosomal markers. The IgG effect was independent of other particle-associated antigens or serum factors. Lysosome/phagosome attachment was also quantified biochemically with a cytosol-dependent scintillation proximity assay. Interactions were enhanced significantly in reactions containing cytosol from mouse macrophages that had been exposed to IgG-coated beads, indicating that IgG signaling modulates the cytosolic-targeting machinery. Similar results were obtained with cytosol from primary human monocytes, human U-937 histiocytic lymphoma cells and from Chinese hamster ovary (CHO) cells transfected with a human IgG (Fcgamma) receptor. IgG-induced activation is shown to affect the actin-dependent tethering/docking stage of the targeting process and to proceed through a pathway involving protein kinase C. These results provide a rare example of an extracellular signal controlling membrane targeting on the level of tethering and docking. We propose that this pathway contributes to the role of antibodies in the protection against microbial infections.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
A cell-free scintillation proximity assay for studies on lysosome-to-phagosome targeting.Sci STKE. 2007 Jun 26;2007(392):pl3. doi: 10.1126/stke.3922007pl3. Sci STKE. 2007. PMID: 17595222
-
Mycobacterium tuberculosis phagosomes exhibit altered calmodulin-dependent signal transduction: contribution to inhibition of phagosome-lysosome fusion and intracellular survival in human macrophages.J Immunol. 2001 Mar 1;166(5):3392-401. doi: 10.4049/jimmunol.166.5.3392. J Immunol. 2001. PMID: 11207296
-
Mycobacterial PknG Targets the Rab7l1 Signaling Pathway To Inhibit Phagosome-Lysosome Fusion.J Immunol. 2018 Sep 1;201(5):1421-1433. doi: 10.4049/jimmunol.1800530. Epub 2018 Jul 23. J Immunol. 2018. PMID: 30037848
-
Better Together: Current Insights Into Phagosome-Lysosome Fusion.Front Immunol. 2021 Feb 25;12:636078. doi: 10.3389/fimmu.2021.636078. eCollection 2021. Front Immunol. 2021. PMID: 33717183 Free PMC article. Review.
-
How nascent phagosomes mature to become phagolysosomes.Trends Immunol. 2012 Aug;33(8):397-405. doi: 10.1016/j.it.2012.03.003. Epub 2012 May 3. Trends Immunol. 2012. PMID: 22560866 Review.
Cited by
-
Antibodies Induced by Lipoarabinomannan in Bovines: Characterization and Effects on the Interaction between Mycobacterium Avium Subsp. Paratuberculosis and Macrophages In Vitro.Vet Med Int. 2011;2011:258479. doi: 10.4061/2011/258479. Epub 2011 Jun 8. Vet Med Int. 2011. PMID: 21772964 Free PMC article.
-
Multisystem disorder associated with a pathogenic variant in CLCN7 in the absence of osteopetrosis.Mol Genet Genomic Med. 2024 Jul;12(7):e2494. doi: 10.1002/mgg3.2494. Mol Genet Genomic Med. 2024. PMID: 39056574 Free PMC article.
-
Noncanonical autophagy is required for type I interferon secretion in response to DNA-immune complexes.Immunity. 2012 Dec 14;37(6):986-997. doi: 10.1016/j.immuni.2012.09.014. Epub 2012 Dec 6. Immunity. 2012. PMID: 23219390 Free PMC article.
-
Dengue virus compartmentalization during antibody-enhanced infection.Sci Rep. 2017 Jan 13;7:40923. doi: 10.1038/srep40923. Sci Rep. 2017. PMID: 28084461 Free PMC article.
-
Determinants of natural immunity against tuberculosis in an endemic setting: factors operating at the level of macrophage-Mycobacterium tuberculosis interaction.Clin Exp Immunol. 2008 Mar;151(3):414-22. doi: 10.1111/j.1365-2249.2007.03585.x. Clin Exp Immunol. 2008. PMID: 18234054 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
