

Genes affecting cell competition in Drosophila
- PMID: 17110495
- PMCID: PMC1800612
- DOI: 10.1534/genetics.106.061929
Genes affecting cell competition in Drosophila
Abstract
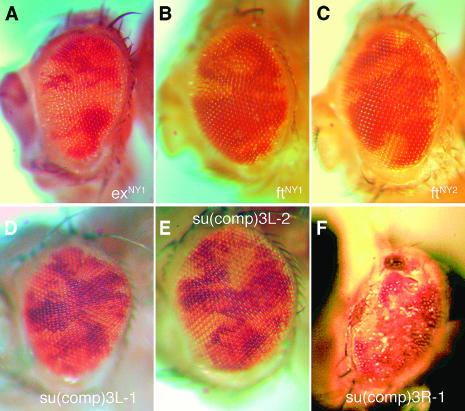
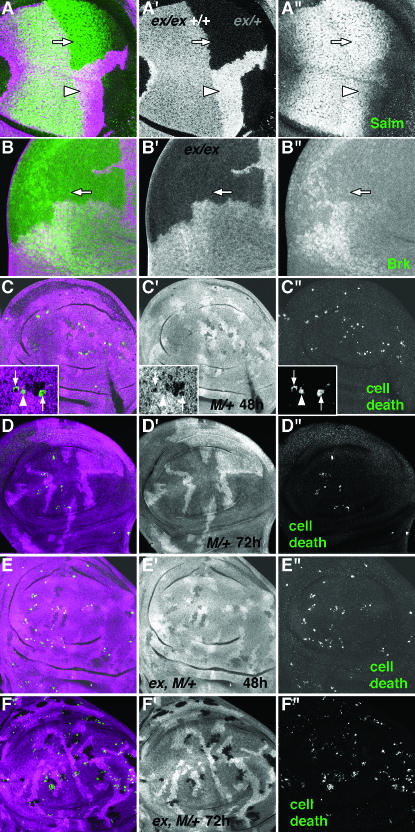
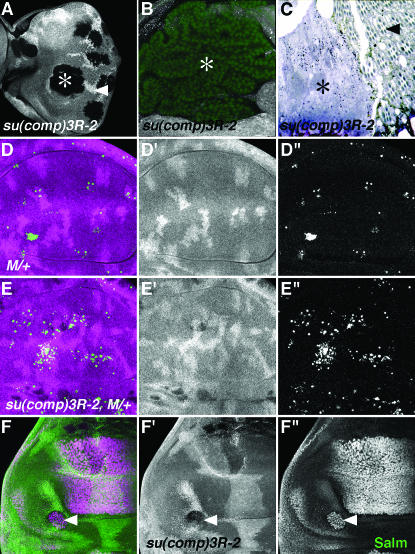
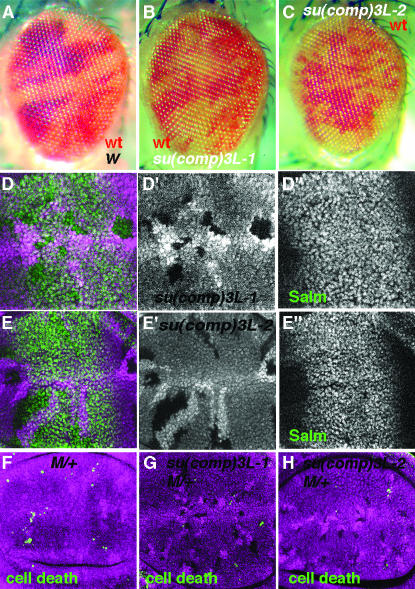
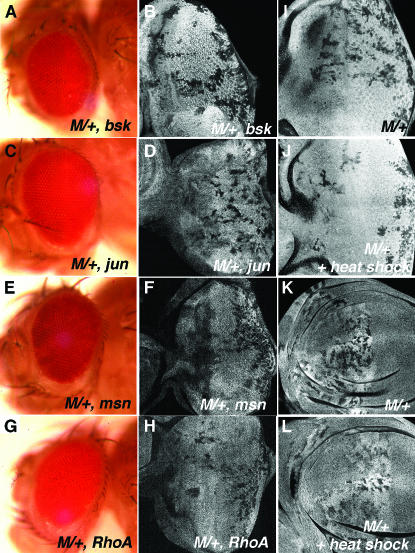
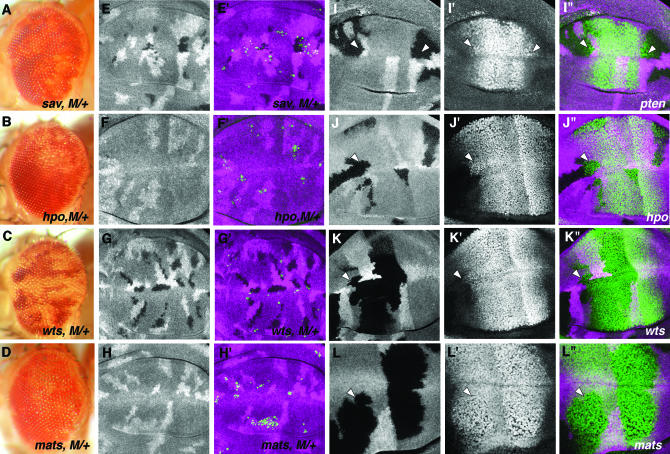
Cell competition is a homeostatic mechanism that regulates the size attained by growing tissues. We performed an unbiased genetic screen for mutations that permit the survival of cells being competed due to haplo-insufficiency for RpL36. Mutations that protect RpL36 heterozygous clones include the tumor suppressors expanded, hippo, salvador, mats, and warts, which are members of the Warts pathway, the tumor suppressor fat, and a novel tumor-suppressor mutation. Other hyperplastic or neoplastic mutations did not rescue RpL36 heterozygous clones. Most mutations that rescue cell competition elevated Dpp-signaling activity, and the Dsmurf mutation that elevates Dpp signaling was also hyperplastic and rescued. Two nonlethal, nonhyperplastic mutations prevent the apoptosis of Minute heterozygous cells and suggest an apoptosis pathway for cell competition . In addition to rescuing RpL36 heterozygous cells, mutations in Warts pathway genes were supercompetitors that could eliminate wild-type cells nearby. The findings show that differences in Warts pathway activity can lead to competition and implicate the Warts pathway, certain other tumor suppressors, and novel cell death components in cell competition, in addition to the Dpp pathway implicated by previous studies. We suggest that cell competition might occur during tumor development in mammals.
Figures




Similar articles
-
Differences in levels of the transmembrane protein Crumbs can influence cell survival at clonal boundaries.Dev Biol. 2012 Aug 15;368(2):358-69. doi: 10.1016/j.ydbio.2012.06.001. Epub 2012 Jun 7. Dev Biol. 2012. PMID: 22683826 Free PMC article.
-
The ligand Sas and its receptor PTP10D drive tumour-suppressive cell competition.Nature. 2017 Feb 9;542(7640):246-250. doi: 10.1038/nature21033. Epub 2017 Jan 16. Nature. 2017. PMID: 28092921
-
A genetic screen in Drosophila for metastatic behavior.Science. 2003 Nov 14;302(5648):1227-31. doi: 10.1126/science.1088474. Epub 2003 Oct 9. Science. 2003. PMID: 14551319
-
Regulation of imaginal disc growth by tumor-suppressor genes in Drosophila.Annu Rev Genet. 2006;40:335-61. doi: 10.1146/annurev.genet.39.073003.100738. Annu Rev Genet. 2006. PMID: 16872256 Review.
-
Cell competition and its implications for development and cancer.J Genet Genomics. 2011 Oct 20;38(10):483-95. doi: 10.1016/j.jgg.2011.09.006. Epub 2011 Sep 21. J Genet Genomics. 2011. PMID: 22035869 Free PMC article. Review.
Cited by
-
Cell competition removes segmental aneuploid cells from Drosophila imaginal disc-derived tissues based on ribosomal protein gene dose.Elife. 2021 Apr 13;10:e61172. doi: 10.7554/eLife.61172. Elife. 2021. PMID: 33847264 Free PMC article.
-
Cell competition: how to eliminate your neighbours.Development. 2014 Mar;141(5):988-1000. doi: 10.1242/dev.079129. Development. 2014. PMID: 24550108 Free PMC article. Review.
-
Coupling of Hedgehog and Hippo pathways promotes stem cell maintenance by stimulating proliferation.J Cell Biol. 2014 May 12;205(3):325-38. doi: 10.1083/jcb.201309141. Epub 2014 May 5. J Cell Biol. 2014. PMID: 24798736 Free PMC article.
-
Myc Function in Drosophila.Genes Cancer. 2010 Jun 1;1(6):542-546. doi: 10.1177/1947601910377490. Genes Cancer. 2010. PMID: 21072325 Free PMC article.
-
Drosophila follicle stem cells are regulated by proliferation and niche adhesion as well as mitochondria and ROS.Nat Commun. 2012 Apr 3;3:769. doi: 10.1038/ncomms1765. Nat Commun. 2012. PMID: 22473013 Free PMC article.
References
-
- Adler, P. N., J. Charlton, K. H. Jones and J. Liu, 1994. The cold-sensitive period for frizzled in the development of wing hair polarity ends prior to the start of hair morphogenesis. Mech. Dev. 46: 101–107. - PubMed
-
- Adler, P. N., J. Charlton and J. Liu, 1998. Mutations in the cadherin superfamily member gene dachsous cause a tissue polarity phenotype by altering frizzled signaling. Development 125: 959–968. - PubMed
-
- Berger, J., T. Suzuki, K. A. Senti, J. Stubbs, G. Schaffner et al., 2001. Genetic mapping with SNP markers in Drosophila. Nat. Genet. 29: 475–481. - PubMed
-
- Bilder, D., and N. Perrimon, 2000. Localization of apical epithelial determinants by the basolateral PDZ protein Scribble. Nature 403: 676–680. - PubMed
-
- Blaumueller, C. M., and M. Mlodzik, 2000. The Drosophila tumor suppressor expanded regulates growth, apoptosis, and patterning during development. Mech. Dev. 92: 251–262. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
