

The cooperative response of synaptotagmin I C2A. A hypothesis for a Ca2+-driven molecular hammer
- PMID: 17114221
- PMCID: PMC1783886
- DOI: 10.1529/biophysj.106.087197
The cooperative response of synaptotagmin I C2A. A hypothesis for a Ca2+-driven molecular hammer
Abstract
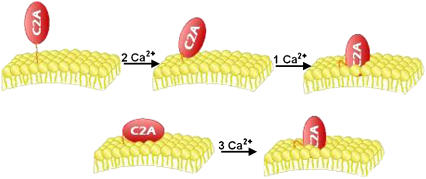
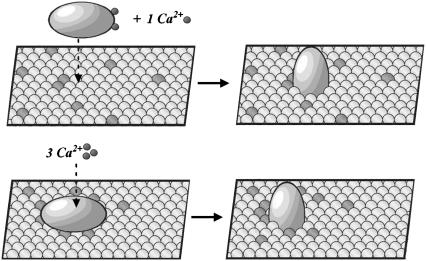
In the current understanding of exocytosis at the nerve terminal, the C2 domain of synaptotagmin (C2A) is presumed to bind Ca2+ and the membrane in a stepwise fashion: cation then membrane as cation increases the affinity of protein for membrane. Fluorescence spectroscopy data were gathered over a variety of lipid and Ca2+ concentrations, enabling the rigorous application of microscopic binding models derived from partition functions to differentiate between Ca2+ and phosphatidylserine contributions to binding. The data presented here are in variance with previously published models, which were based on the Hill approximation. Rather, the data are consistent with two forms of cooperativity that modulate the responsiveness of C2A: in Ca2+ binding to a network of three cation sites and in interaction with the membrane surface. We suggest synaptotagmin I C2A is preassociated with the synaptic vesicle membrane or nerve terminal. In this state, upon Ca2+ influx the protein will bind the three Ca2+ ions immediately and with high cooperativity. Thus, membrane association creates a high-affinity Ca2+ switch that is the basis for the role of synaptotagmin I in Ca2+-regulated exocytosis. Based on this model, we discuss the implications of protein-induced phosphatidylserine demixing to the exocytotic process.
Figures





References
-
- Fernandez-Chacon, R., A. Königstorfer, S. H. Gerber, J. Garcia, M. F. Matos, C. F. Stevens, N. Brose, J. Rizo, C. Rosenmund, and T. C. Sudhof. 2001. Synaptotagmin I functions as a calcium regulator of release probability. Nature. 410:41–49. - PubMed
-
- Shin, O.-H., J.-S. Rhee, J. Tang, S. Sugita, C. Rosenmund, and T. C. Sudhof. 2003. Sr2+ binding to the Ca2+ binding site of the synaptotagmin 1 C2B domain triggers fast exocytosis without stimulating SNARE interactions. Neuron. 37:99–109. - PubMed
-
- Sudhof, T. C. 2004. The synaptic vesicle cycle. Annu. Rev. Neurosci. 27:509–547. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
