

doi: 10.1101/gad.1475106.
Epub 2006 Nov 17.
Wnt/beta-catenin signaling regulates vertebrate limb regeneration
Affiliations
- PMID: 17114576
- PMCID: PMC1686599
- DOI: 10.1101/gad.1475106
Item in Clipboard
Wnt/beta-catenin signaling regulates vertebrate limb regeneration
Genes Dev.
.
Abstract
The cellular and molecular bases allowing tissue regeneration are not well understood. By performing gain- and loss-of-function experiments of specific members of the Wnt pathway during appendage regeneration, we demonstrate that this pathway is not only necessary for regeneration to occur, but it is also able to promote regeneration in axolotl, Xenopus, and zebrafish. Furthermore, we show that changes in the spatiotemporal distribution of beta-catenin in the developing chick embryo elicit apical ectodermal ridge and limb regeneration in an organism previously thought not to regenerate. Our studies may provide valuable insights toward a better understanding of adult tissue regeneration.
Figures
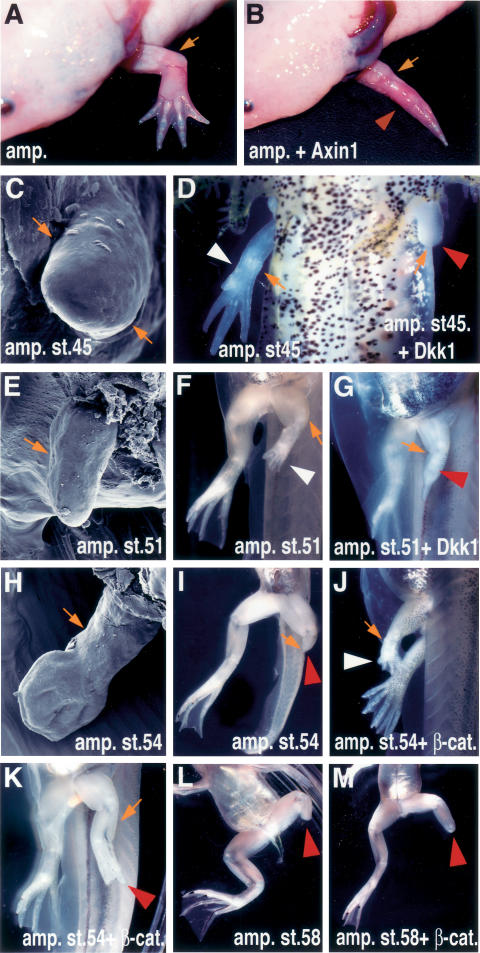
Wnt/β-catenin signaling is required for limb regeneration in axolotls and Xenopus. (A–D) Gross morphology and scanning electron microscope (SEM) images of adult (A,B) and larvae (C,D) axolotls. (A) A fully regenerated wild-type adult axolotl forelimb develops 2 mo after amputation at the elbow level. (B) Microinjection of Ad-Axin1 after forelimb amputation prevented regeneration of the distal elements (red arrowhead). (C) SEM image of an axolotl larvae limb at the stage at which amputation was performed (stage 45). (D) Ventral view of axolotl larvae limbs 1 mo after amputation and virus injection. The right amputated limb regenerated normally (reader’s left side, white arrowhead), while the left amputated limb, injected with Ad-Dkk1, did not regenerate (reader’s right side, red arrowhead). (E–M) Gross morphology and SEM images of Xenopus larvae hindlimbs. (E) SEM image of the Xenopus larvae hindlimb at the amputated stage 51. (F) Ventral view of the control limb 21 d after amputation. The limb regenerated (white arrowhead) and formed the autopod. (G) Ventral view of the limb 21 d after amputation and injection of Ad-Dkk1. Dkk1 inhibited regeneration (red arrowhead), and prevented the formation of the more distal limb elements. (H) SEM image of Xenopus larvae hindlimb at the amputated stage (54). (I) Ventral view of the limb 21 d after amputation. The limb did not regenerate when amputated at stage 54 (red arrowhead). Lateral (J) and ventral (K) views of the limb 21 d after amputation and injection of Ad-CA-β-catenin. β-Catenin was able to regenerate the distal limb elements (white arrowhead, J), or partially restore regeneration of distal structures (red arrowhead, K). (L) Ventral view of a limb 21 d after amputation at the knee level at stage 58, showing lack of regeneration. (M) Ventral view of a limb 21 d after amputation and injection of Ad-CA-β-catenin at the knee level at stage 58, showing lack of regeneration. A small outgrowth is observed. In larvae shown in F–M, amputation with or without virus injection was done to the left limb while the right limb was left to develop normally. Orange arrows indicate the amputation level, and white and red arrowheads indicate regenerated and regeneration-defective limbs, respectively.
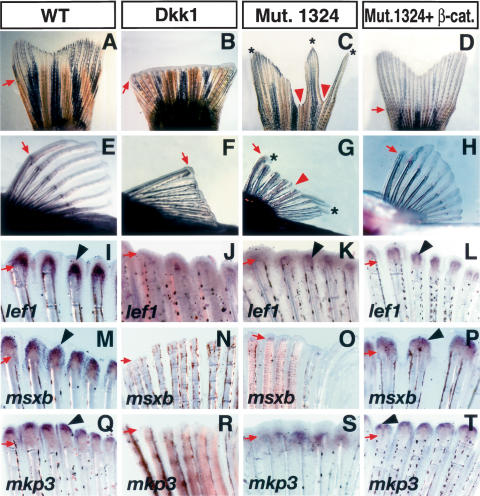
Wnt/β-catenin is necessary and enhances fin regeneration in zebrafish. (A–H) Gross morphology of the caudal fin (A–D) and pectoral fin (E–H) of adult zebrafish 7–10 d after amputation with or without virus injection. The caudal fin (A) and pectoral fin (E) regenerated normally in wild-type zebrafish after amputation. Ad-Dkk1 injection after amputation inhibited regeneration in both the caudal fin (B) and pectoral fin (F), demonstrating the necessity of Wnt/β-catenin signaling for fin regeneration. The #1324 mutant fish caudal fin (C) and pectoral fin (G) showed defective regeneration, and only certain areas of the fin showed regeneration (asterisks). Injection of Ad-CA-β-catenin restored fin regeneration in the #1324 mutant. Both the caudal fin (D) and pectoral fin (H) regenerated similarly to that of wild-type fins. (I–T) In situ hybridization of lef1 (I–L), msxb (M–P), and mkp3 (Q–T) in pectoral fins 2 d after amputation with or without virus injection. In the wild-type fin, lef1 (I), msxb (M), and mkp3 (Q) are strongly expressed. In Ad-Dkk1 injected fins, expression of lef1 (J), msxb (N), and mkp3 (R) is barely detectable. In #1324 mutant fins, expression of lef1 (K), msxb (O), and mkp3 (S) is significantly down-regulated when compared with control fins. Injection of Ad-CA-β-catenin restored expression of lef1 (L), msxb (P), and mkp3 (T) in the #1324 mutant fin. Arrows indicate the amputation level. Red arrowheads and asterisks in C and G indicate defects in regeneration and regenerated tissue in the #1324 mutant. Arrowheads in I–T indicate gene expression.
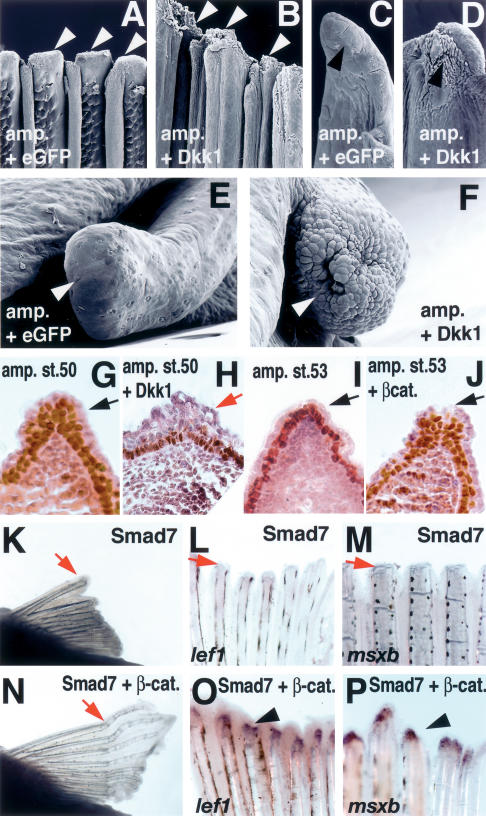
Wnt and BMP signaling controls AEC formation during fin/limb regeneration. (A–F) SEM images of zebrafish pectoral fin (A–D) and axolotl larval limb (E,F) injected with Ad-eGFP (A,C,E) and Ad-Dkk1 (B,D,F). (A,C) Surface of the control pectoral fin with Ad-eGFP at 24 h post-amputation shows the characteristic smooth ectoderm (arrowheads). (B,D) Injection of Ad-Dkk1 caused thickening and alterations in the shape of ectodermal cells (arrowheads). C and D are close-ups of A and B, respectively, showing a single ray. (E) Axolotl larvae limb at 2 d post-amputation and Ad-eGFP-injected shows a smooth surface covering the amputation site (arrowhead). (F) Ad-Dkk1-injected larvae limb shows a rough and disorganized surface at the amputation site 2 d post-amputation (arrowhead). (G–J) Difference in ectoderm stratification during Xenopus hindlimb regeneration as revealed by immunoreactivity for p63 (brown color). Sections were counterstained with hematoxylin-eosin. (G) A Xenopus hindlimb, amputated at stage 50, formed stratified ectoderm 7 d after amputation (arrow). (H) Amputation of a stage 50 hindlimb, followed by injection of Ad-Dkk1, resulted in a single layer of ectoderm (red arrow). (I) Amputation at stage 53 resulted in the formation of a single layer of ectoderm after 15 d (red arrow). (J) Injection of Ad-CA-β-catenin to stage 53 amputated hindlimb induced ectoderm stratification after 15 d, similar to the one observed in G (arrow). (K–P) BMP/Smad signaling is necessary for pectoral fin regeneration and CA-β-catenin can restore BMP-inhibited regeneration. Gross morphology of pectoral fin (K,N) at 7 d post-amputation and expression of lef1 (L,O) and msxb (M,P) at 2 d post-amputation Ad-Smad7-injected (K–M) and coinjection of Ad-Smad7 + Ad-CA-β-catenin (N–P). Inhibition of BMP/Smad signaling by overexpression of Smad7 inhibited fin regeneration (K), lef1 (L), and msxb expression (M). Coinjection of Ad-CA-β-catenin restored the defects in regeneration caused by Smad7 (N) and lef1 (O) msxb expression (P).
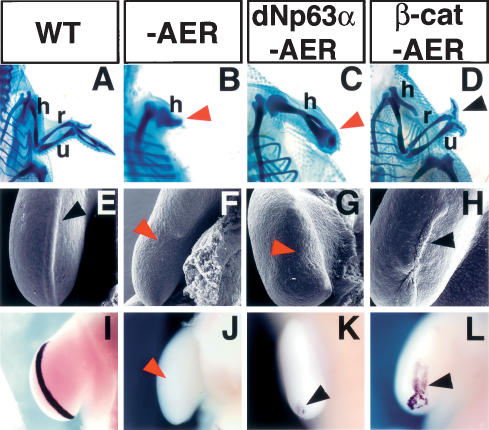
β-Catenin induces AER and limb regeneration in chick embryos. (A–D) Cartilage staining of the chick forelimb at 10 d post-fertilization. (A) Wild-type chick limb showing the stylopod (humerus), zeugopod (radius and ulna), and autopod elements. (B,C) Removal of the AER at stage 20/21 prevented limb outgrowth and resulted in a truncated limb independent of dNp63α overexpression. (D) Ad-CA-β-catenin injection prior to AER removal restored limb development and induced the formation of all the stylopod, zeugopod elements, and two digits (black arrowhead). Red arrowheads in B and C indicate truncated limbs. (E–L) SEM images (E–H) and fgf8 expression (I–L) of the limb ectoderm 30 h after AER removal. The control limb shows the typical thickened AER ectoderm (black arrowhead, E) associated with fgf8 expression (I). Absence of distal thickened ectoderm after AER removal (red arrowhead, F) results in the down-regulation of fgf8 expression (red arrowhead, J). Overexpression of dNp63α did not rescue AER (red arrowhead, G) or fgf8 expression (residual expression is observed at the posterior of the developing limb bud) (black arrowhead, K). Infection of limb ectoderm cells with Ad-CA-β-catenin prior to AER removal regenerated the AER (black arrowhead, H) and elicited fgf8 expression (black arrowhead, L). (h) Humerus; (r) radius; (u) ulna.
References
-
- Ahn, K., Mishina, Y., Hanks, M.C., Behringer, R.R., Crenshaw, E.B., III BMPR-IA signaling is required for the formation of the apical ectodermal ridge and dorsal–ventral patterning of the limb. Development. 2001;128:4449–4461. - PubMed
-
- Akimenko, M.A., Mari-Beffa, M., Becerra, J., Geraudie, J. Old questions, new tools, and some answers to the mystery of fin regeneration. Dev. Dyn. 2003;226:190–201. - PubMed
-
- Bakkers, J., Hild, M., Kramer, C., Furutani-Seiki, M., Hammerschmidt, M. Zebrafish ΔNp63 is a direct target of Bmp signaling and encodes a transcriptional repressor blocking neural specification in the ventral ectoderm. Dev. Cell. 2002;2:617–627. - PubMed
-
- Bode, H.R. Head regeneration in Hydra. Dev. Dyn. 2003;226:225–236. - PubMed
-
- Brockes, J.P., Kumar, A. Plasticity and reprogramming of differentiated cells in amphibian regeneration. Nat. Rev. Mol. Cell Biol. 2002;3:566–574. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases