

The S. cerevisiae Rrm3p DNA helicase moves with the replication fork and affects replication of all yeast chromosomes
- PMID: 17114583
- PMCID: PMC1635146
- DOI: 10.1101/gad.1478906
The S. cerevisiae Rrm3p DNA helicase moves with the replication fork and affects replication of all yeast chromosomes
Abstract
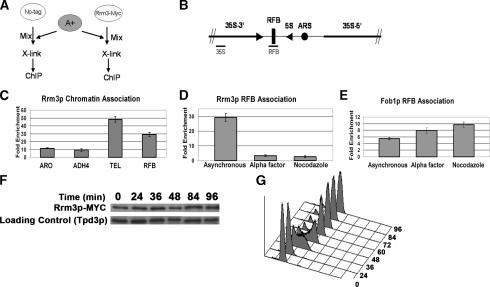
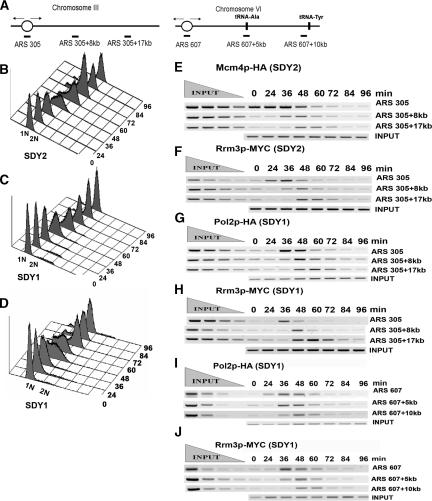
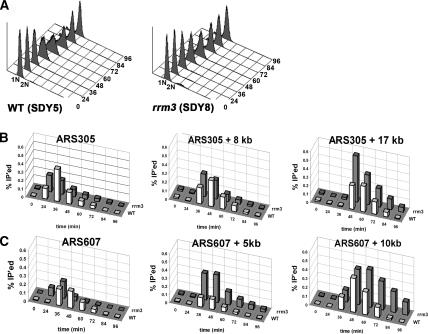
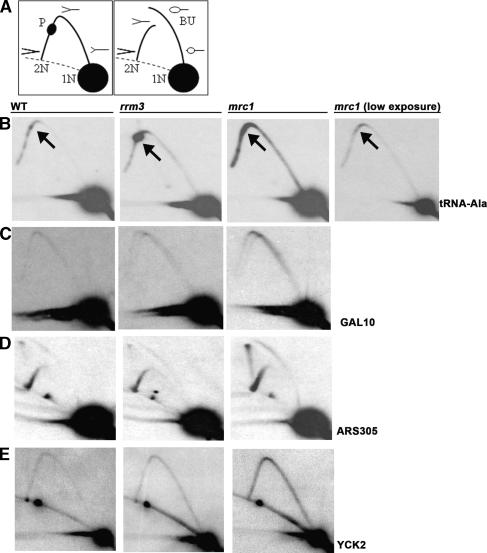
The Saccharomyces cerevisiae DNA helicase Rrm3p is needed for normal fork progression through >1000 discrete sites scattered throughout the genome. Here we show that replication of all yeast chromosomes was markedly delayed in rrm3 cells. Delayed replication was seen even in a region that lacks any predicted Rrm3p-dependent sites. Based on the pattern of replication intermediates in two-dimensional gels, the rate of fork movement in rrm3 cells appeared similar to wild-type except at known Rrm3p-dependent sites. These data suggest that although Rrm3p has a global role in DNA replication, its activity is needed only or primarily at specific, difficult-to-replicate sites. By the criterion of chromatin immunoprecipitation, Rrm3p was associated with both Rrm3p-dependent and -independent sites, and moved with the replication fork through both. In addition, Rrm3p interacted with Pol2p, the catalytic subunit of DNA polymerase epsilon, in vivo. Thus, rather than being recruited to its sites of action when replication forks stall at these sites, Rrm3p is likely a component of the replication fork apparatus.
Figures



References
-
- Alcasabas, A., Osborn, A., Bachant, J., Hu, F., Werler, P., Bousset, K., Furuya, K., Diffley, J., Carr, A., Elledge, S. Mrc1 transduces signals of DNA replication stress to activate Rad53. Nat. Cell Biol. 2001;3:958–965. - PubMed
-
- Aparicio, O.M., Weinstein, D.M., Bell, S.P. Components and dynamics of DNA replication complexes in S. cerevisiae: Redistribution of MCM proteins and Cdc45p during S phase. Cell. 1997;91:59–69. - PubMed
-
- Barry, J., Alberts, B. A role for two DNA helicases in the replication of T4 bacteriophage DNA. J. Biol. Chem. 1994;269:33063–33068. - PubMed
-
- Bedinger, P., Hochstrasser, M., Jongeneel, C., Alberts, B. Properties of the T4 bacteriophage DNA replication apparatus: The T4 dda DNA helicase is required to pass a bound RNA polymerase molecule. Cell. 1983;34:115–123. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases