

Review
doi: 10.4161/rna.3.1.2495.
Epub 2006 Jan 12.
A-to-I RNA editing and human disease
Affiliations
- PMID: 17114938
- PMCID: PMC2947206
- DOI: 10.4161/rna.3.1.2495
Item in Clipboard
Review
A-to-I RNA editing and human disease
RNA Biol.
2006 Jan-Mar.
Abstract
The post-transcriptional modification of mammalian transcripts by A-to-I RNA editing has been recognized as an important mechanism for the generation of molecular diversity and also regulates protein function through recoding of genomic information. As the molecular players of editing are characterized and an increasing number of genes become identified that are subject to A-to-I modification, the potential impact of editing on the etiology or progression of human diseases is realized. Here we review the recent knowledge on where disturbances in A-to-I RNA editing have been correlated with human disease phenotypes.
Figures
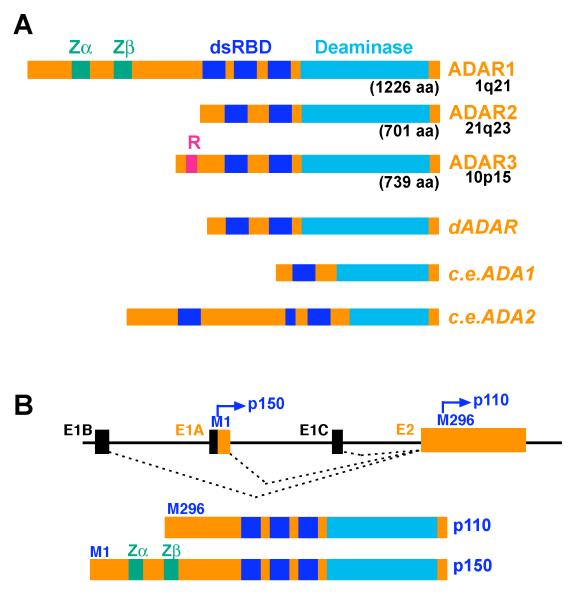
A) The structure of vertebrate and invertebrate ADAR gene family members. Indicated are two Z-DNA binding domains (ADAR1), the dsRNA binding (dsRBDs), the arginine-rich (R-domain), ssRNA binding domain (ADAR3) and the deaminase domain. For human ADAR1-3 amino acid lengths and chromosomal localization are shown. The structure of Drosophila dADAR and two C. elegans c.e.ADA1 and c.e.ADA2 are shown. Drosophila dADAR is very similar to mammalian ADAR2. c.e.ADA1 contains a single copy of a poorly conserved dsRBD and a conserved deaminase domain, whereas c.e.ADA2 contains three copies of dsRBDs and a deaminase domain that is rather divergent from any known ADAR genes. B) Two ADAR1 translation products. Exons E1A (IFN/dsRNA-inducible) and E1B and E1C (constitutive) are spliced to exon 2 at precisely the same junction. Exon 1A contains the Met initiation codon for the 1226-aa ORF specifying the IFN/dsRNA-inducible p150 protein. Exon 1B and 1C do not contain an AUG initiation codon. The second AUG initiation codon located in exon 2 initiates translation of the 931-aa encoding the constitutively expressed ADAR1 p110 protein.

Total of 32 mutations (wild-type, green; mutation, red) associated with DSH have been identified in Japanese and Chinese populations -. Indicated are Zα and Zβ domains (brown overline), three dsRNA binding domains dsRBD1-3 (yellow filled box), and nine stretches of core deaminase subdomains, highly conserved in all three mammalian ADAR gene family members ADAR1-3 (red box). Two methionines (M1 and M296) used for alternative translation initiation of p150 and p110, respectively, are highlighted in blue.

The specific reduction in glutamate receptor GluR-2 Q/R site editing in ALS motor neurons leads to excessive influx of Ca2+-ions through unedited glutamate gated channels (right panel). In concert with other molecular events that increase glutamate toxicity (glutamate transporter deficiency) motor neurons a subjected to increased stress that leads to accumulation of radical oxygen species (ROS), protein degradation dysfunction, neurofilament entanglement and eventually, neuronal death. N= nucleus; GT=glutamate transporter

A; Genomic DNA and aligned amino acid sequence of the 5-HT2CR intracellular loop II region. Five RNA editing sites detected (A-E sites) within exon 5 are shown in red. A combination of A-to-I RNA editing events at these sites (detected as A-to-G cDNA sequence change) result in various amino acid sequence alterations (open box). B; The seven-transmembrane receptor 5-HT2CR (dark gray cylinders with loops) is shown embedded within the lipid bilayer (lighter yellow in the background). Seven amino acids (white characters in brown circles) in the intracellular loop II region important for G-protein coupling are depicted together with substitutions brought on by editing (red characters in black circles) at these locations. The unedited protein isoform is denoted as “INI” indicating the amino acids specified by the unedited codons. The fully edited isoform transforming “INI” to “VGV” profoundly reduces 5-HT potency, G-protein coupling and agonist binding, thereby affecting downstream signaling pathways. Among the edited 5-HT2CR isoforms, the asparagine to glycine change at position 158 causes the most profound alteration in function and is therefore shown with slightly larger size. C; Formation of a RNA fold back structure involving exon 5 (lightly shaded)/intron 5 of the human 5-HT2CR gene transcript is necessary for site-selective editing at the A-E sites by ADARs as well as for intronic F site editing. Editing of the A and B sites is preferentially catalyzed by ADAR1 and the D site by ADAR2. The E and C sites may be edited efficiently by both ADAR1 and ADAR2. Among three alternative splicing donor sites (GU1 to GU3 in green), only GU2 leads to the mature mRNA that produces functional, full-length 5-HT2CR protein. The editing pattern modulates splice site selection.
References
-
- Johnson JM, et al. Genome-wide survey of human alternative pre-mRNA splicing with exon junction microarrays. Science. 2003;302:2141–4. - PubMed
-
- Keegan LP, Gallo A, O’Connell MA. The many roles of an RNA editor. Nat Rev Genet. 2001;2:869–78. - PubMed
-
- Gott JM, Emeson RB. Functions and mechanisms of RNA editing. Annu Rev Genet. 2000;34:499–531. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources