

Abr and Bcr, two homologous Rac GTPase-activating proteins, control multiple cellular functions of murine macrophages
- PMID: 17116687
- PMCID: PMC1800684
- DOI: 10.1128/MCB.00756-06
Abr and Bcr, two homologous Rac GTPase-activating proteins, control multiple cellular functions of murine macrophages
Abstract
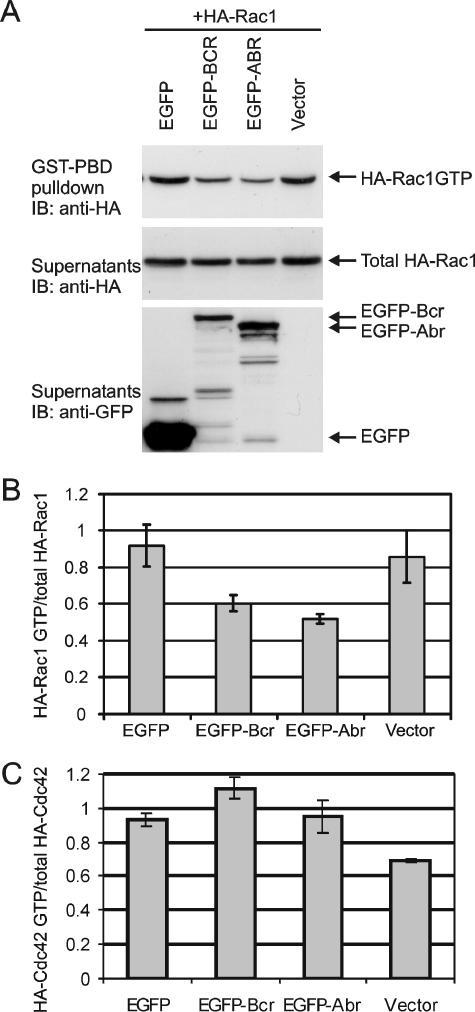
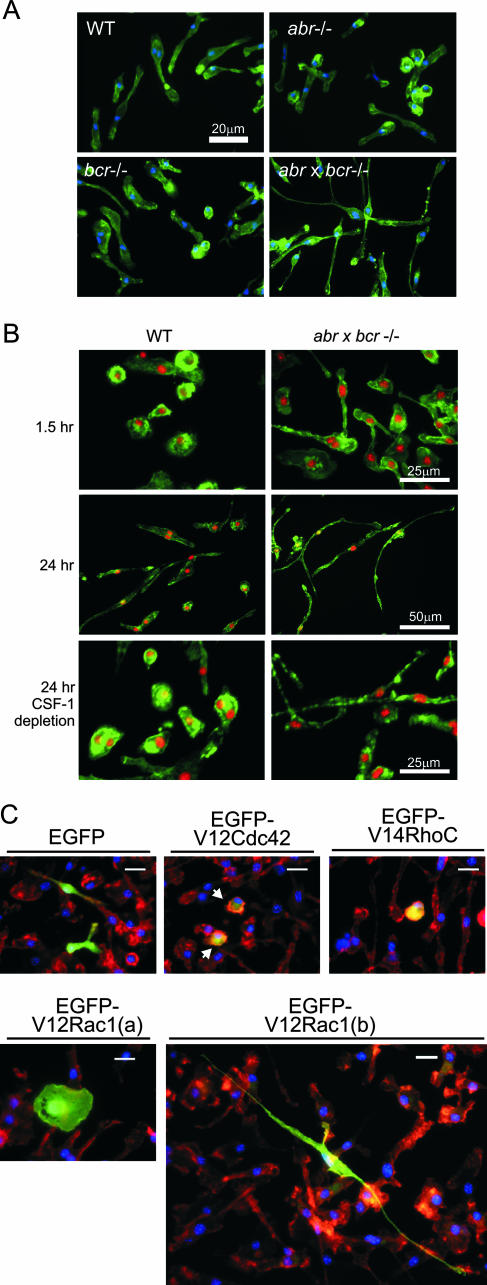
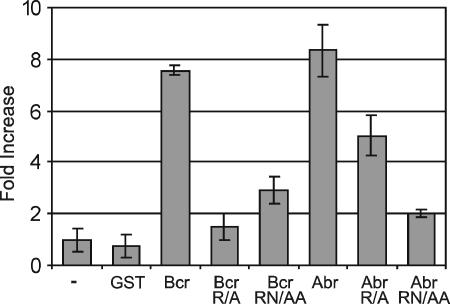
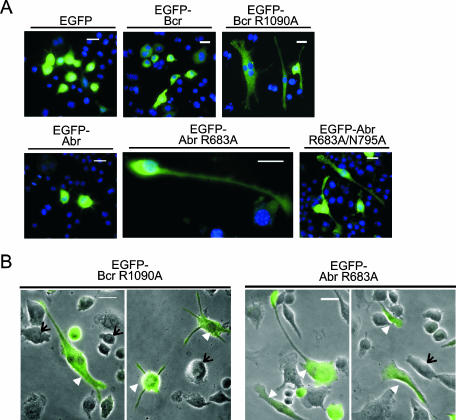
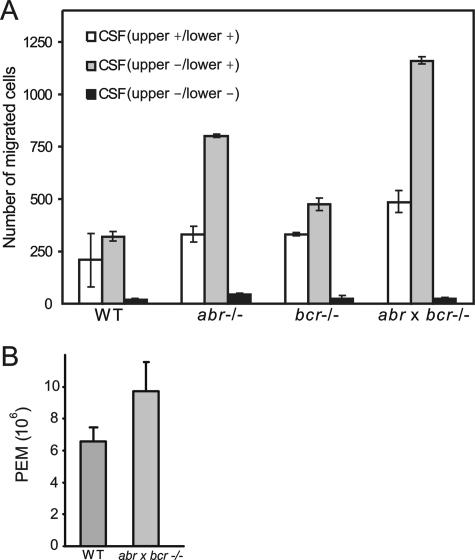
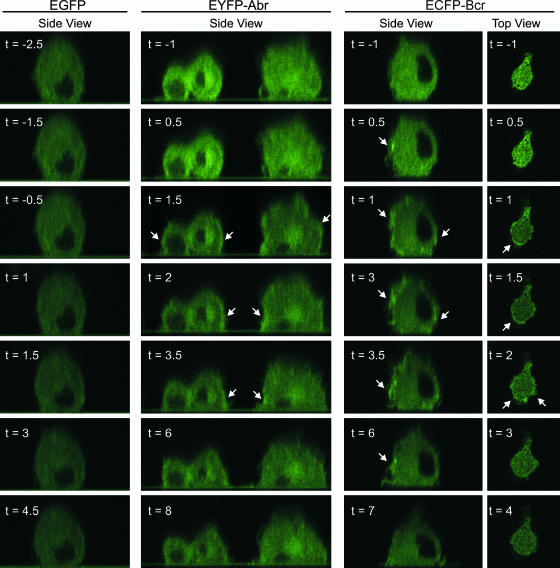
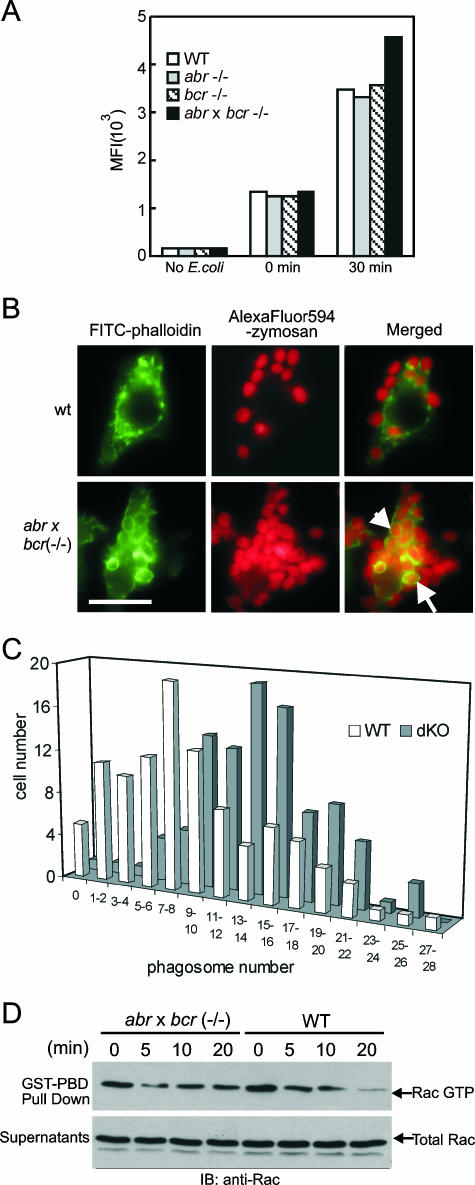
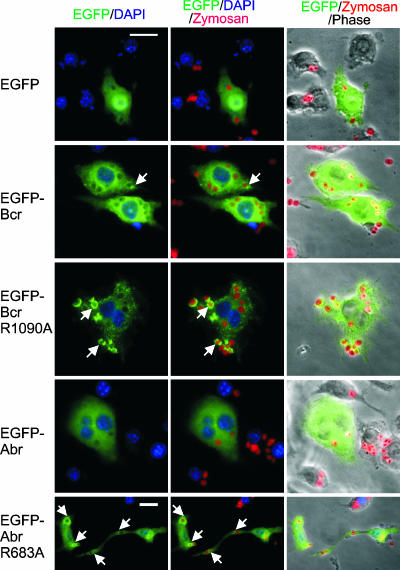
Small GTPases of the Rho family are key regulators of phagocytic leukocyte function. Abr and Bcr are homologous, multidomain proteins. Their C-terminal domain has GTPase-activating protein (GAP) activity that, in vitro, is specific for Rac and Cdc42. To address the in vivo relevance of these entire proteins, of which little is known, the current study examined the effect of the genetic ablation of Abr and Bcr in murine macrophages. The concomitant loss of Abr and Bcr induced multiple alterations of macrophage cellular behavior known to be under the control of Rac. Macrophages lacking both Abr and Bcr exhibited an atypical, elongated morphology that was reproduced by the ectopic expression of GAP domain mutant Abr and Bcr in a macrophage cell line and of constitutively active Rac in primary macrophages. A robust increase in colony-stimulating factor 1 (CSF-1)-directed motility was observed in macrophages deficient for both proteins and, in response to CSF-1 stimulation, Abr and Bcr transiently translocated to the plasma membrane. Phagocytosis of opsonized particles was also increased in macrophages lacking both proteins and correlated with sustained Rac activation. Bcr and Abr GAP mutant proteins localized around phagosomes and induced distinct phagocytic cup formation. These results identify Abr and Bcr as the only GAPs to date that specifically negatively regulate Rac function in vivo in primary macrophages.
Figures
Similar articles
-
Lack of bcr and abr promotes hypoxia-induced pulmonary hypertension in mice.PLoS One. 2012;7(11):e49756. doi: 10.1371/journal.pone.0049756. Epub 2012 Nov 12. PLoS One. 2012. PMID: 23152932 Free PMC article.
-
Bcr and Abr cooperate in negatively regulating acute inflammatory responses.Mol Cell Biol. 2009 Nov;29(21):5742-50. doi: 10.1128/MCB.00357-09. Epub 2009 Aug 24. Mol Cell Biol. 2009. PMID: 19703997 Free PMC article.
-
Activity of the Bcr GTPase-activating domain is regulated through direct protein/protein interaction with the Rho guanine nucleotide dissociation inhibitor.J Biol Chem. 2008 Feb 8;283(6):3023-3030. doi: 10.1074/jbc.M705513200. Epub 2007 Dec 10. J Biol Chem. 2008. PMID: 18070886
-
The Rho GTPases in macrophage motility and chemotaxis.Cell Adhes Commun. 1998;6(2-3):237-45. doi: 10.3109/15419069809004479. Cell Adhes Commun. 1998. PMID: 9823474 Review.
-
Dendritic cells: new roles for Cdc42 and Rac in antigen uptake?Curr Biol. 2000 Oct 19;10(20):R739-41. doi: 10.1016/s0960-9822(00)00736-3. Curr Biol. 2000. PMID: 11069097 Review.
Cited by
-
Differential Transcriptome Networks between IDO1-Knockout and Wild-Type Mice in Brain Microglia and Macrophages.PLoS One. 2016 Jun 17;11(6):e0157727. doi: 10.1371/journal.pone.0157727. eCollection 2016. PLoS One. 2016. PMID: 27314674 Free PMC article.
-
Abr, a Rho-regulating protein, modulates osteoclastogenesis by enhancing lamellipodia formation by interacting with poly(ADP-ribose) glycohydrolase.Mol Biol Rep. 2023 Sep;50(9):7557-7569. doi: 10.1007/s11033-023-08690-0. Epub 2023 Jul 28. Mol Biol Rep. 2023. PMID: 37507586
-
Src homology 2 domain containing protein 5 (SH2D5) binds the breakpoint cluster region protein, BCR, and regulates levels of Rac1-GTP.J Biol Chem. 2014 Dec 19;289(51):35397-408. doi: 10.1074/jbc.M114.615112. Epub 2014 Oct 20. J Biol Chem. 2014. PMID: 25331951 Free PMC article.
-
Control of synapse development and plasticity by Rho GTPase regulatory proteins.Prog Neurobiol. 2011 Jul;94(2):133-48. doi: 10.1016/j.pneurobio.2011.04.011. Epub 2011 Apr 22. Prog Neurobiol. 2011. PMID: 21530608 Free PMC article. Review.
-
Identification and selected reaction monitoring (SRM) quantification of endocytosis factors associated with Numb.Mol Cell Proteomics. 2013 Feb;12(2):499-514. doi: 10.1074/mcp.M112.020768. Epub 2012 Dec 4. Mol Cell Proteomics. 2013. PMID: 23211419 Free PMC article.
References
-
- Aghazadeh, B., K. Zhu, T. J. Kubiseski, G. A. Liu, T. Pawson, Y. Zheng, and M. K. Rosen. 1998. Structure and mutagenesis of the Dbl homology domain. Nat. Struct. Biol. 5:1098-1107. - PubMed
-
- Allen, W. E., G. E. Jones, J. W. Pollard, and A. J. Ridley. 1997. Rho, Rac and Cdc42 regulate actin organization and cell adhesion in macrophages. J. Cell Sci. 110:707-720. - PubMed
-
- Benard, V., and G. M. Bokoch. 2002. Assay of Cdc42, Rac, and Rho GTPase activation by affinity methods. Methods Enzymol. 345:349-359. - PubMed
-
- Bernards, A. 2003. GAPs galore! A survey of putative Ras superfamily GTPase activating proteins in man and Drosophila. Biochim. Biophys. Acta 1603:47-82. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous