

Calmodulin potentiates G beta gamma activation of phospholipase C-beta3
- PMID: 17118346
- PMCID: PMC1866284
- DOI: 10.1016/j.bcp.2006.10.004
Calmodulin potentiates G beta gamma activation of phospholipase C-beta3
Abstract
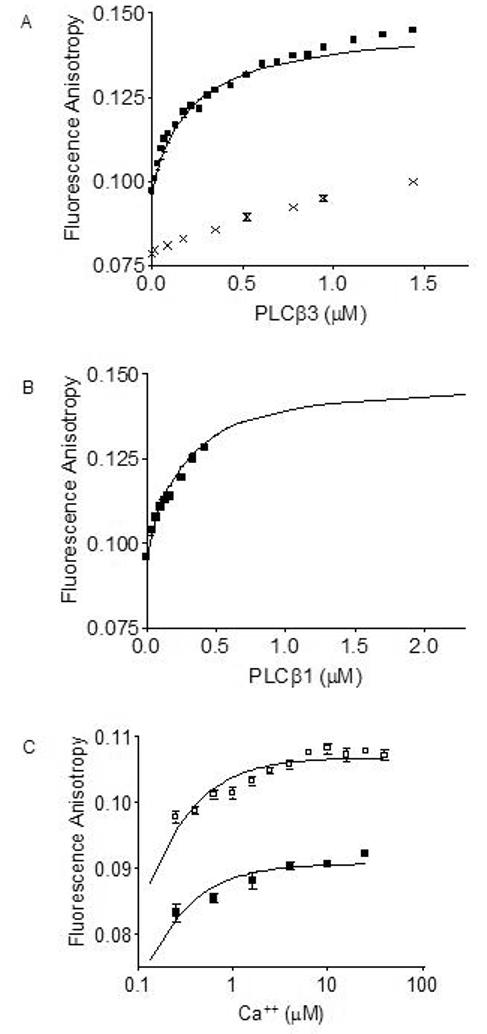
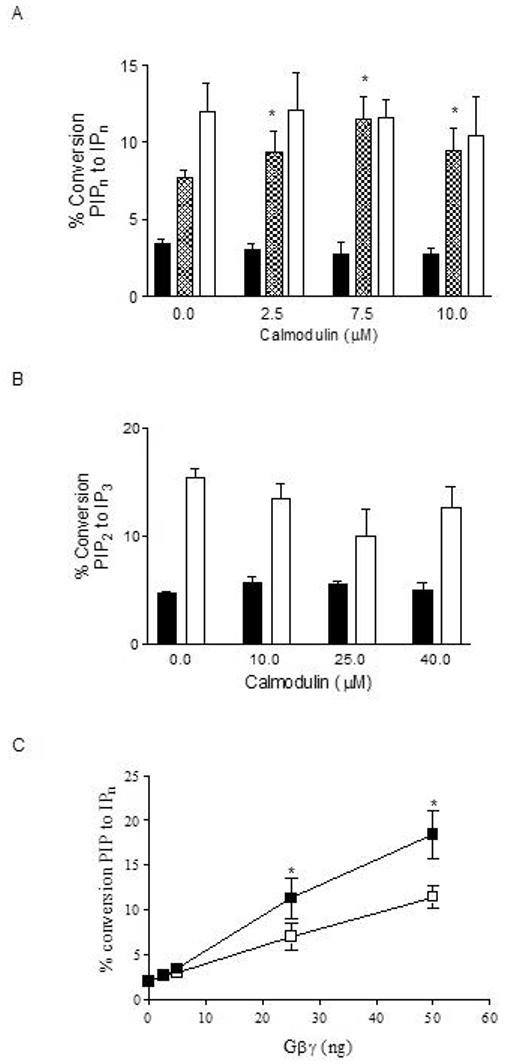
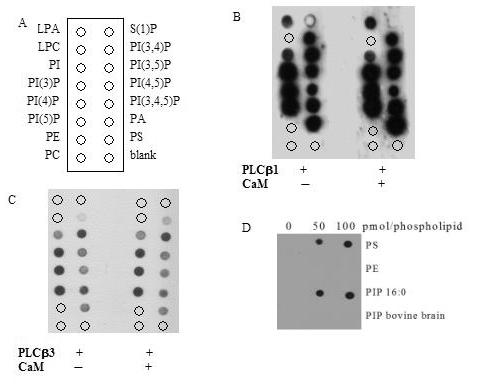
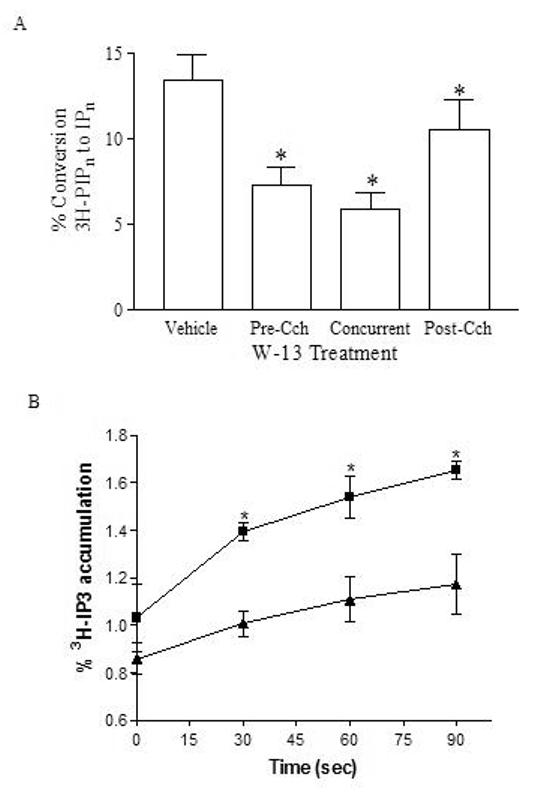
Phospholipase C-beta (PLC-beta) isozymes (EC 3.1.4.11) hydrolyze the membrane phospholipid phosphatidylinositol-4,5-bisphosphate to generate intracellular second messenger signaling molecules inositol-1,4,5-trisphosphate (IP3) and diacylglycerol (DAG) in response to receptor activation and other cellular stimuli. PLCbeta1 and PLCbeta3 isozymes were previously demonstrated to bind the calcium-sensitive molecule calmodulin [McCullar JS, Larsen SA, Millimaki RA, Filtz TM. Calmodulin is a phospholipase C-{beta} interacting protein. J Biol Chem 2003;278(36):33708-13]. We have now shown through fluorescence anisotropy that calmodulin/PLCbeta3 affinities increase with increasing calcium in a physiologically relevant concentration range. The bimolecular affinity constants for calmodulin interaction with PLCbeta1 or PLCbeta3 were estimated as 260 and 200 nM, respectively, from fluorescence anisotropy data. There was no effect of calmodulin on basal or G alpha q-stimulated catalytic activity for either isozyme. However, the interaction between calmodulin and PLCbeta3 leads to potentiation of activation by the G-protein beta gamma dimer in an in vitro assay. 1321N1 cells treated with calmodulin inhibitors concurrent with and post-stimulation of muscarinic receptors significantly reduced [3H]PIP hydrolysis. Together these data are suggestive of cooperative role for calmodulin in the G-protein beta gamma dimer-stimulated activity of PLCbeta3.
Figures
References
-
- McCullar JS, Larsen SA, Millimaki RA, Filtz TM. Calmodulin Is a Phospholipase C-{beta} Interacting Protein. J. Biol. Chem. 2003;278(36):33708–33713. - PubMed
-
- Rebecchi MJ, Pentyala SN. Structure, Function, and Control of Phosphoinositide-Specific Phospholipase C. Physiological Reviews. 2000;80:1291–1335. - PubMed
-
- Song C, Hu C-D, Masago M, Kariya K-i, Yamawaki-Kataoka Y, Shibatohge M, Wu D, Satoh T, Kataoka T. Regulation of a Novel Human Phospholipase C, PLCepsilon, through Membrane Targeting by Ras. J. Biol. Chem. 2001;276(4):2752–2757. - PubMed
-
- Saunders CM, Larman MG, Parrington J, Cox LJ, Royse J, Blayney LM, Swann K, Lai FA. PLC{zeta}: a sperm-specific trigger of Ca2+ oscillations in eggs and embryo development. Development. 2002;129(15):3533–3544. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
