

The fusiform face area: a cortical region specialized for the perception of faces
- PMID: 17118927
- PMCID: PMC1857737
- DOI: 10.1098/rstb.2006.1934
The fusiform face area: a cortical region specialized for the perception of faces
Abstract
Faces are among the most important visual stimuli we perceive, informing us not only about a person's identity, but also about their mood, sex, age and direction of gaze. The ability to extract this information within a fraction of a second of viewing a face is important for normal social interactions and has probably played a critical role in the survival of our primate ancestors. Considerable evidence from behavioural, neuropsychological and neurophysiological investigations supports the hypothesis that humans have specialized cognitive and neural mechanisms dedicated to the perception of faces (the face-specificity hypothesis). Here, we review the literature on a region of the human brain that appears to play a key role in face perception, known as the fusiform face area (FFA). Section 1 outlines the theoretical background for much of this work. The face-specificity hypothesis falls squarely on one side of a longstanding debate in the fields of cognitive science and cognitive neuroscience concerning the extent to which the mind/brain is composed of: (i) special-purpose ('domain-specific') mechanisms, each dedicated to processing a specific kind of information (e.g. faces, according to the face-specificity hypothesis), versus (ii) general-purpose ('domain-general') mechanisms, each capable of operating on any kind of information. Face perception has long served both as one of the prime candidates of a domain-specific process and as a key target for attack by proponents of domain-general theories of brain and mind. Section 2 briefly reviews the prior literature on face perception from behaviour and neurophysiology. This work supports the face-specificity hypothesis and argues against its domain-general alternatives (the individuation hypothesis, the expertise hypothesis and others). Section 3 outlines the more recent evidence on this debate from brain imaging, focusing particularly on the FFA. We review the evidence that the FFA is selectively engaged in face perception, by addressing (and rebutting) five of the most widely discussed alternatives to this hypothesis. In section 4, we consider recent findings that are beginning to provide clues into the computations conducted in the FFA and the nature of the representations the FFA extracts from faces. We argue that the FFA is engaged both in detecting faces and in extracting the necessary perceptual information to recognize them, and that the properties of the FFA mirror previously identified behavioural signatures of face-specific processing (e.g. the face-inversion effect). Section 5 asks how the computations and representations in the FFA differ from those occurring in other nearby regions of cortex that respond strongly to faces and objects. The evidence indicates clear functional dissociations between these regions, demonstrating that the FFA shows not only functional specificity but also area specificity. We end by speculating in section 6 on some of the broader questions raised by current research on the FFA, including the developmental origins of this region and the question of whether faces are unique versus whether similarly specialized mechanisms also exist for other domains of high-level perception and cognition.
Figures
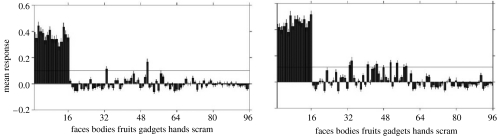
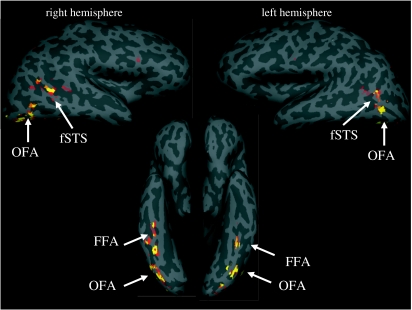
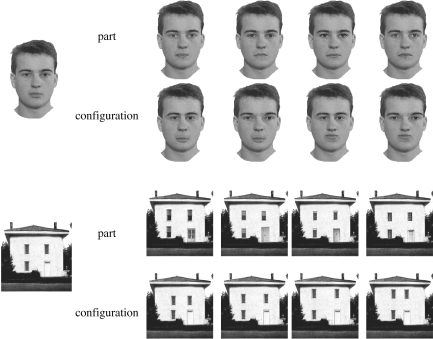
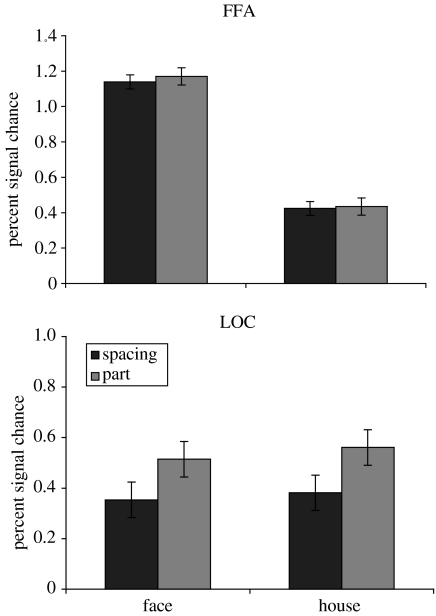
References
-
- Aguirre G.K, Singh R, D'Esposito M. Stimulus inversion and the responses of face and object-sensitive cortical areas. Neuroreport. 1999;10:189–194. - PubMed
-
- Allison T, Ginter H, McCarthy G, Nobre A.C, Puce A, Luby M, Spencer D.D. Face recognition in human extrastriate cortex. J. Neurophysiol. 1994;71:821–825. - PubMed
-
- Allison T, Puce A, Spencer D.D, McCarthy G. Electrophysiological studies of human face perception. I: potentials generated in occipitotemporal cortex by face and non-face stimuli. Cereb. Cortex. 1999;9:415–430. doi:10.1093/cercor/9.5.415 - DOI - PubMed
-
- Andrews T.J, Ewbank M.P. Distinct representations for facial identity and changeable aspects of faces in the human temporal lobe. Neuroimage. 2004;23:905–913. doi:10.1016/j.neuroimage.2004.07.060 - DOI - PubMed
-
- Andrews T.J, Schluppeck D. Neural responses to Mooney images reveal a modular representation of faces in human visual cortex. Neuroimage. 2004;21:91–98. doi:10.1016/j.neuroimage.2003.08.023 - DOI - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
