

Behavioural and neurophysiological evidence for face identity and face emotion processing in animals
- PMID: 17118930
- PMCID: PMC1764842
- DOI: 10.1098/rstb.2006.1937
Behavioural and neurophysiological evidence for face identity and face emotion processing in animals
Abstract
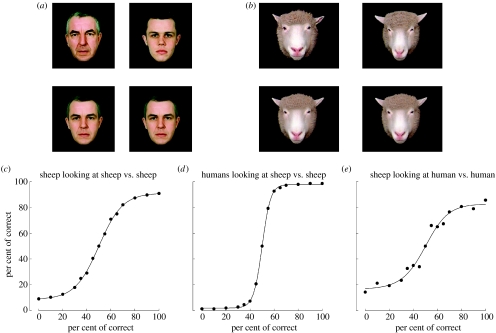
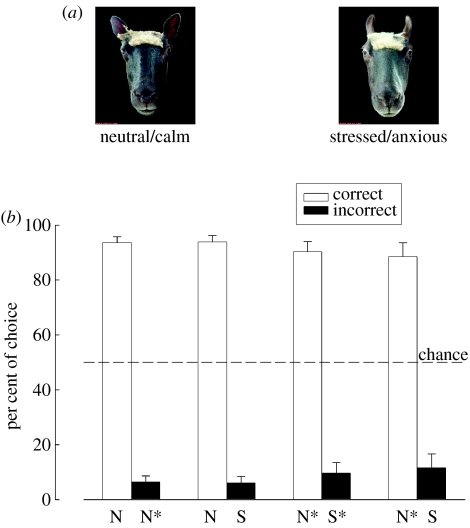
Visual cues from faces provide important social information relating to individual identity, sexual attraction and emotional state. Behavioural and neurophysiological studies on both monkeys and sheep have shown that specialized skills and neural systems for processing these complex cues to guide behaviour have evolved in a number of mammals and are not present exclusively in humans. Indeed, there are remarkable similarities in the ways that faces are processed by the brain in humans and other mammalian species. While human studies with brain imaging and gross neurophysiological recording approaches have revealed global aspects of the face-processing network, they cannot investigate how information is encoded by specific neural networks. Single neuron electrophysiological recording approaches in both monkeys and sheep have, however, provided some insights into the neural encoding principles involved and, particularly, the presence of a remarkable degree of high-level encoding even at the level of a specific face. Recent developments that allow simultaneous recordings to be made from many hundreds of individual neurons are also beginning to reveal evidence for global aspects of a population-based code. This review will summarize what we have learned so far from these animal-based studies about the way the mammalian brain processes the faces and the emotions they can communicate, as well as associated capacities such as how identity and emotion cues are dissociated and how face imagery might be generated. It will also try to highlight what questions and advances in knowledge still challenge us in order to provide a complete understanding of just how brain networks perform this complex and important social recognition task.
Figures





References
-
- Aggleton J.P, Young A.W. The enigma of the amygdala: on its contribution to human emotion. In: Lane R.D, Nadel L, editors. Cognitive neuroscience of emotion. Oxford University Press; New York, NY: 2000. pp. 106–128.
-
- Baker C.I, Keysers C, Jellema T, Wicker B, Perrett D.I. Neuronal representation of disappearing and hidden objects in temporal cortex of the macaque. Exp. Brain. Res. 2001;140:375–381. doi:10.1007/s002210100828 - DOI - PubMed
-
- Bauer H.R, Philip M. Facial and vocal individual recognition in the common chimpanzee. Psychol. Rec. 1983;33:161–170.
-
- Baylis G.C, Rolls E.T, Leonard C.M. Selectivity between faces in the responses of a population of neurons in the cortex in the superior temporal sulcus of the monkey. Brain. Res. 1985;342:91–102. doi:10.1016/0006-8993(85)91356-3 - DOI - PubMed
-
- Bekoff M. Animal reflections. Nature. 2002;419:255. doi:10.1038/419255a - DOI - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources