

Mechanisms of regulation of CXCR4/SDF-1 (CXCL12)-dependent migration and homing in multiple myeloma
- PMID: 17119115
- PMCID: PMC1852222
- DOI: 10.1182/blood-2006-07-035857
Mechanisms of regulation of CXCR4/SDF-1 (CXCL12)-dependent migration and homing in multiple myeloma
Abstract
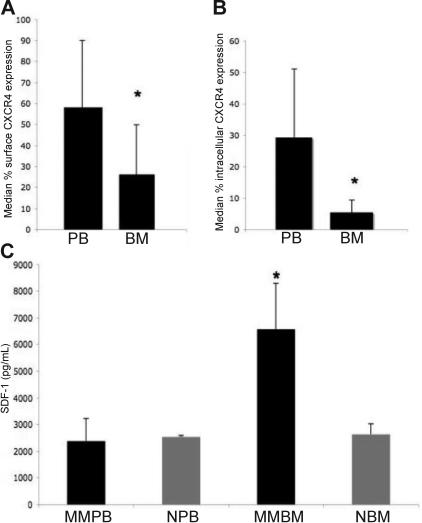
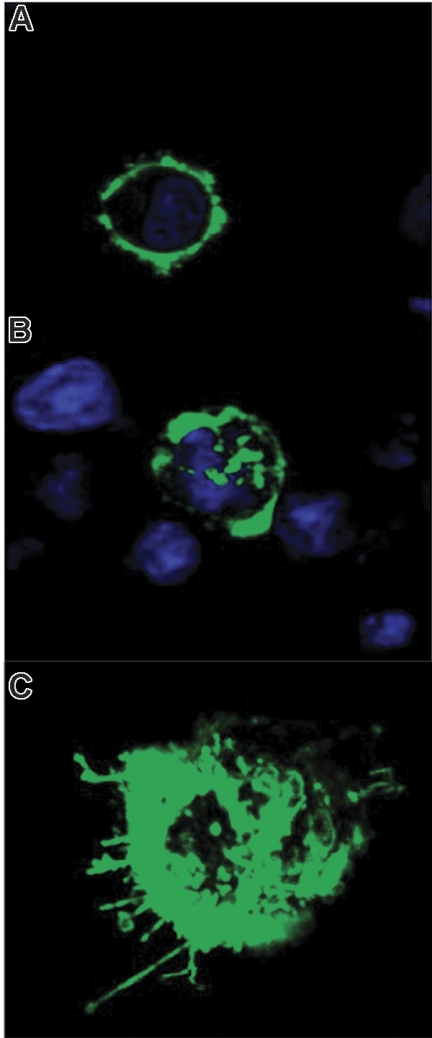
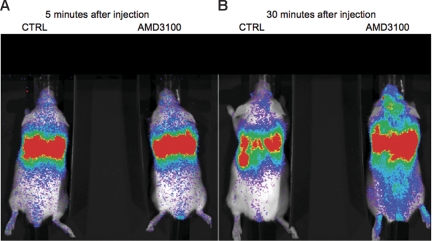
The mechanisms by which multiple myeloma (MM) cells migrate and home to the bone marrow are not well understood. In this study, we sought to determine the effect of the chemokine SDF-1 (CXCL12) and its receptor CXCR4 on the migration and homing of MM cells. We demonstrated that CXCR4 is differentially expressed at high levels in the peripheral blood and is down-regulated in the bone marrow in response to high levels of SDF-1. SDF-1 induced motility, internalization, and cytoskeletal rearrangement in MM cells evidenced by confocal microscopy. The specific CXCR4 inhibitor AMD3100 and the anti-CXCR4 antibody MAB171 inhibited the migration of MM cells in vitro. CXCR4 knockdown experiments demonstrated that SDF-1-dependent migration was regulated by the P13K and ERK/ MAPK pathways but not by p38 MAPK. In addition, we demonstrated that AMD3100 inhibited the homing of MM cells to the bone marrow niches using in vivo flow cytometry, in vivo confocal microscopy, and whole body bioluminescence imaging. This study, therefore, demonstrates that SDF-1/CXCR4 is a critical regulator of MM homing and that it provides the framework for inhibitors of this pathway to be used in future clinical trials to abrogate MM trafficking.
Figures




References
-
- Kyle RA, Rajkumar SV. Multiple myeloma. N Engl J Med. 2004;351:1860–1873. - PubMed
-
- Jemal A, Murray T, Ward E, et al. Cancer statistics, 2005. CA Cancer J Clin. 2005;55:10–30. - PubMed
-
- Billadeau D, Van Ness B, Kimlinger T, et al. Clonal circulating cells are common in plasma cell proliferative disorders: a comparison of monoclonal gammopathy of undetermined significance, smoldering multiple myeloma, and active myeloma. Blood. 1996;88:289–296. - PubMed
-
- Luster AD. Chemokines—chemotactic cytokines that mediate inflammation. N Engl J Med. 1998;338:436–445. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
