

Amelioration of influenza virus pathogenesis in chickens attributed to the enhanced interferon-inducing capacity of a virus with a truncated NS1 gene
- PMID: 17121796
- PMCID: PMC1797581
- DOI: 10.1128/JVI.01667-06
Amelioration of influenza virus pathogenesis in chickens attributed to the enhanced interferon-inducing capacity of a virus with a truncated NS1 gene
Abstract
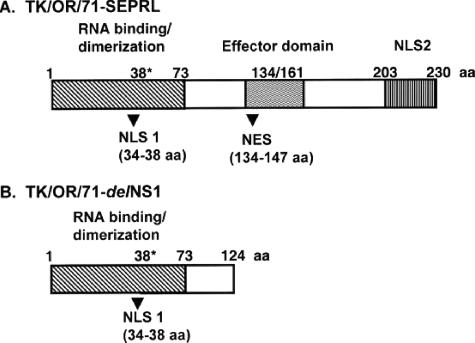
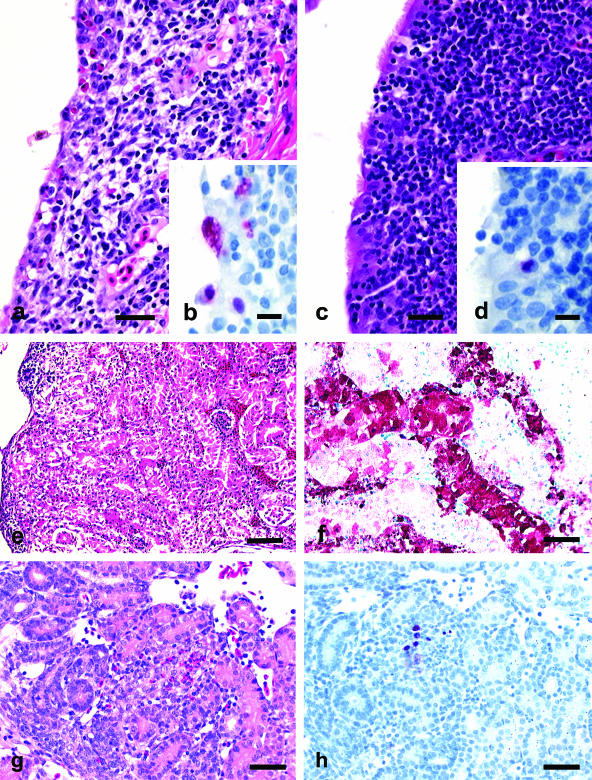
Avian influenza virus (AIV) A/turkey/Oregon/71-SEPRL (TK/OR/71-SEPRL) (H7N3) encodes a full-length NS1 protein and is a weak inducer of interferon (IFN). A variant, TK/OR/71-delNS1 (H7N3), produces a truncated NS1 protein and is a strong inducer of IFN. These otherwise genetically related variants differ 20-fold in their capacities to induce IFN in primary chicken embryo cells but are similar in their sensitivities to the action of IFN. Furthermore, the weak IFN-inducing strain actively suppresses IFN induction in cells that are otherwise programmed to produce it. These phenotypic differences are attributed to the enhanced IFN-inducing capacity that characterizes type A influenza virus strains that produce defective NS1 protein. The pathogenesis of these two variants was evaluated in 1-day-old and 4-week-old chickens. The cell tropisms of both viruses were similar. However, the lesions in chickens produced by the weak IFN inducer were more severe and differed somewhat in character from those observed for the strong IFN inducer. Differences in lesions included the nature of inflammation, the rate of resolution of the infection, and the extent of viral replication and/or virus dissemination. The amelioration of pathogenesis is attributed to the higher levels of IFN produced by the variant encoding the truncated NS1 protein and the antiviral state subsequently induced by that IFN. The high titer of virus observed in kidney tissue ( approximately 10(9) 50% embryo lethal doses/g) from 1-day-old chickens infected intravenously by the weak IFN-inducing strain is attributed to the capacity of chicken kidney cells to activate the hemagglutinin fusion peptide along with their unresponsiveness to inducers of IFN as measured in vitro. Thus, the IFN-inducing capacity of AIV appears to be a significant factor in regulating the pathogenesis, virulence, and viral transmission of AIV in chickens. This suggests that the IFN-inducing and IFN induction suppression phenotypes of AIV should be considered when characterizing strains of influenza virus.
Figures
Similar articles
-
A naturally occurring deletion in its NS gene contributes to the attenuation of an H5N1 swine influenza virus in chickens.J Virol. 2008 Jan;82(1):220-8. doi: 10.1128/JVI.00978-07. Epub 2007 Oct 17. J Virol. 2008. PMID: 17942562 Free PMC article.
-
Prevalence of the C-terminal truncations of NS1 in avian influenza A viruses and effect on virulence and replication of a highly pathogenic H7N1 virus in chickens.Virulence. 2016 Jul 3;7(5):546-57. doi: 10.1080/21505594.2016.1159367. Epub 2016 Mar 16. Virulence. 2016. PMID: 26981790 Free PMC article.
-
A 113-amino-acid truncation at the NS1 C-terminus is a determinant for viral replication of H5N6 avian influenza virus in vitro and in vivo.Vet Microbiol. 2018 Nov;225:6-16. doi: 10.1016/j.vetmic.2018.09.004. Epub 2018 Sep 13. Vet Microbiol. 2018. PMID: 30322535
-
Highly pathogenic avian influenza viruses do not inhibit interferon synthesis in infected chickens but can override the interferon-induced antiviral state.J Virol. 2011 Aug;85(15):7730-41. doi: 10.1128/JVI.00063-11. Epub 2011 May 25. J Virol. 2011. PMID: 21613402 Free PMC article.
-
Modulation of Innate Immune Responses by the Influenza A NS1 and PA-X Proteins.Viruses. 2018 Dec 12;10(12):708. doi: 10.3390/v10120708. Viruses. 2018. PMID: 30545063 Free PMC article. Review.
Cited by
-
Antiviral responses against chicken respiratory infections: Focus on avian influenza virus and infectious bronchitis virus.Cytokine. 2020 Mar;127:154961. doi: 10.1016/j.cyto.2019.154961. Epub 2019 Dec 31. Cytokine. 2020. PMID: 31901597 Free PMC article. Review.
-
High interferon type I responses in the lung, plasma and spleen during highly pathogenic H5N1 infection of chicken.Vet Res. 2011 Jan 11;42(1):6. doi: 10.1186/1297-9716-42-6. Vet Res. 2011. PMID: 21314963 Free PMC article.
-
In vitro analysis of virus particle subpopulations in candidate live-attenuated influenza vaccines distinguishes effective from ineffective vaccines.J Virol. 2010 Nov;84(21):10974-81. doi: 10.1128/JVI.00502-10. Epub 2010 Aug 25. J Virol. 2010. PMID: 20739541 Free PMC article.
-
Influenza Virus: A Master Tactician in Innate Immune Evasion and Novel Therapeutic Interventions.Front Immunol. 2018 Apr 12;9:743. doi: 10.3389/fimmu.2018.00743. eCollection 2018. Front Immunol. 2018. PMID: 29755452 Free PMC article. Review.
-
Upsetting the Balance: When Viruses Manipulate Cell Polarity Control.J Mol Biol. 2018 Sep 28;430(19):3481-3503. doi: 10.1016/j.jmb.2018.04.016. Epub 2018 Apr 20. J Mol Biol. 2018. PMID: 29680664 Free PMC article. Review.
References
-
- Abraham, G. 1979. The effect of ultraviolet radiation on the primary transcription of influenza virus messenger RNAs. Virology 97:177-182. - PubMed
-
- Anonymous. 1999. Guide for the care and use of agricultural animals in agricultural research and teaching. Federation of Animal Science Societies, Savoy, IL.
-
- Ball, L. A. 1979. Induction of 2′5′-oligoadenylate synthetase activity and a new protein by chick interferon. Virology 94:282-296. - PubMed
-
- Beard, C. W., and B. C. Easterday. 1973. A turkey/Oregon/71, an avirulent influenza isolate with hemagglutinin of fowl plague virus. Avian Dis. 17:173-181. - PubMed
-
- Beard, C. W., and D. H. Helfer. 1972. Isolation of two turkey influenza viruses in Oregon. Avian Dis. 16:1133-1136. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Medical
