

Arsenic trioxide affects signal transducer and activator of transcription proteins through alteration of protein tyrosine kinase phosphorylation
- PMID: 17121903
- PMCID: PMC2649745
- DOI: 10.1158/1078-0432.CCR-06-1354
Arsenic trioxide affects signal transducer and activator of transcription proteins through alteration of protein tyrosine kinase phosphorylation
Abstract
Purpose: Arsenic trioxide decreases proliferation of acute myeloid leukemia (AML) cells, but its precise mechanism of action is unknown.
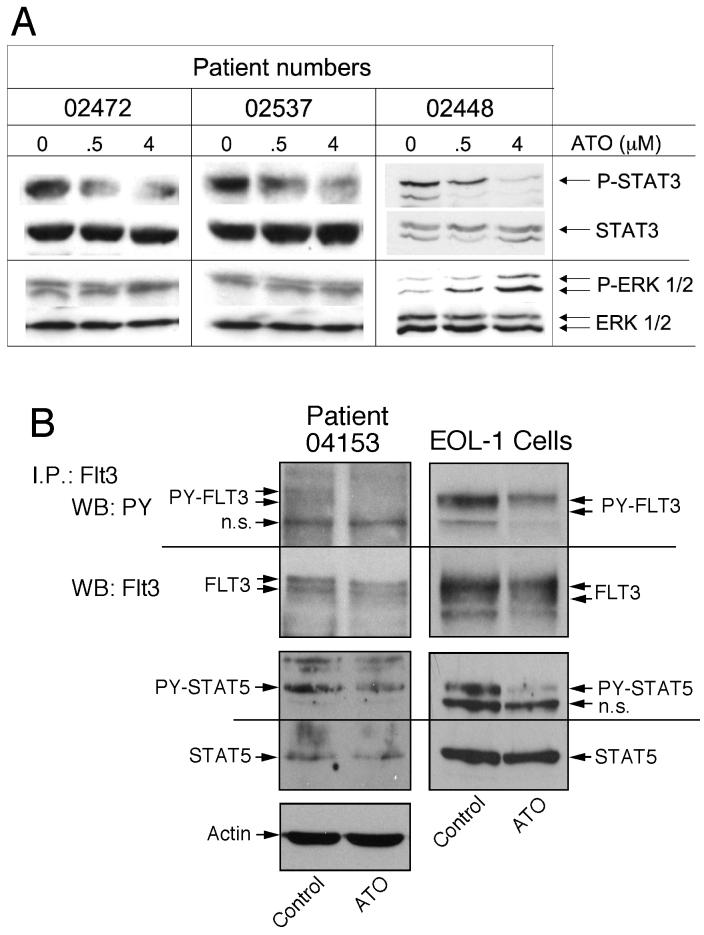
Experimental design: We studied the effect of arsenic trioxide on patient samples and the AML cell line HEL, which, like leukemic blasts from 50% of AML cases, has constitutively activated signal transducer and activator of transcription (STAT) proteins.
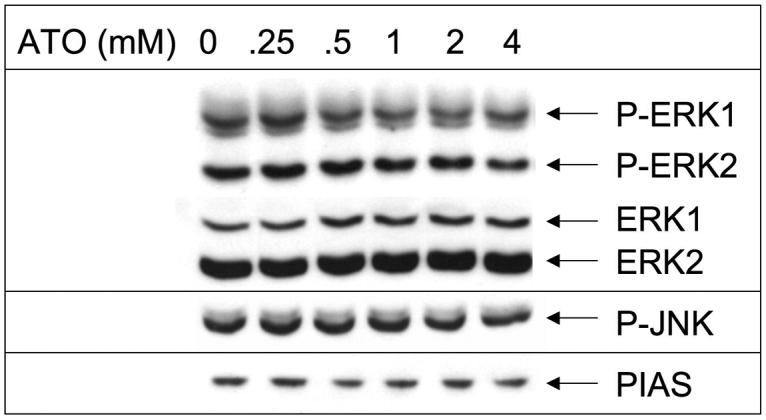
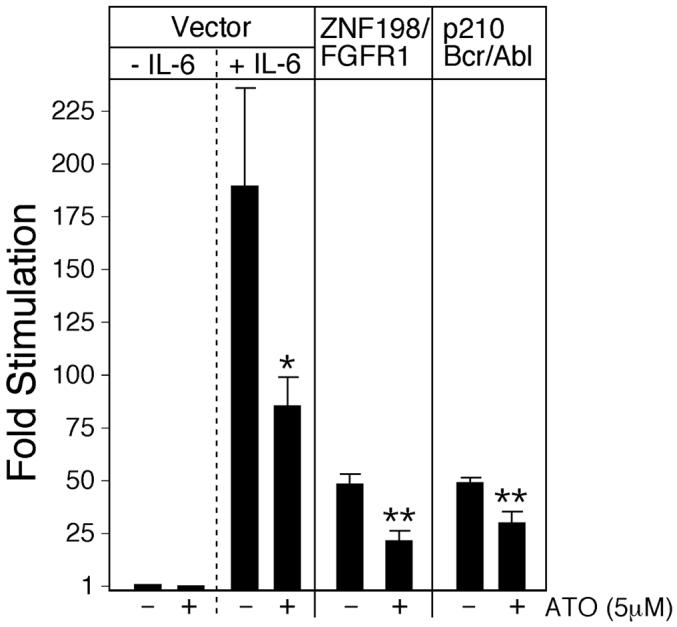
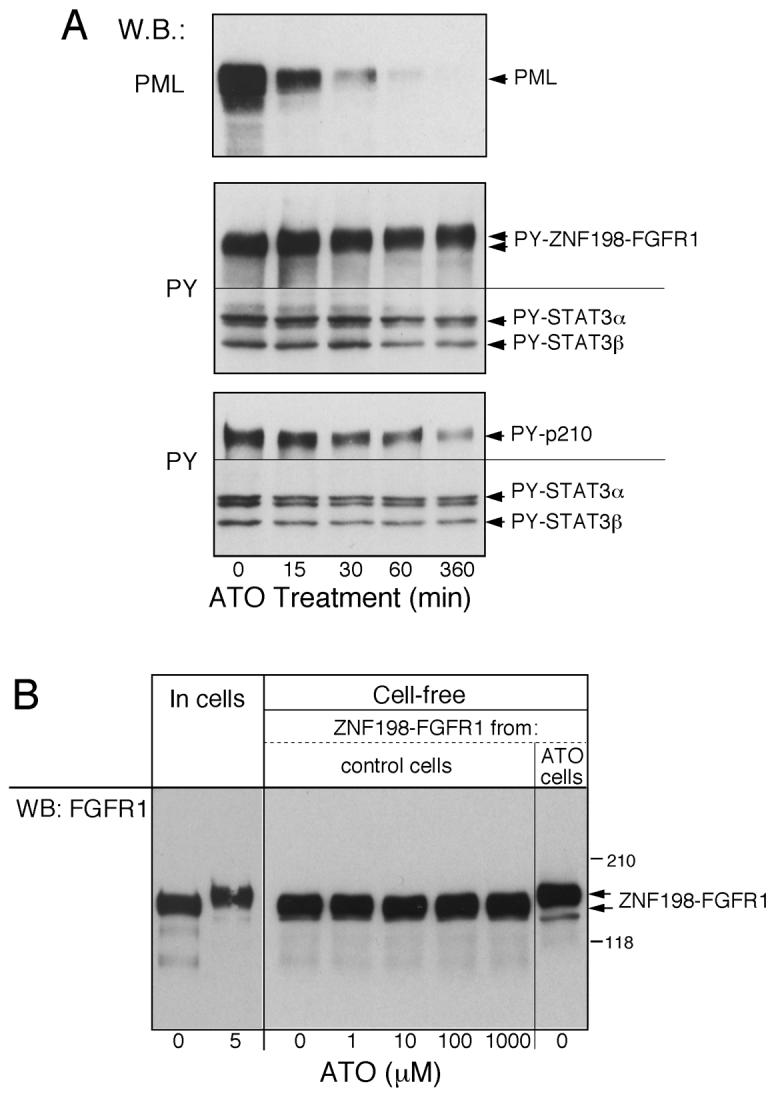
Results: Arsenic trioxide induced mitotic arrest starting at 24 hours and significant cell death at 48 hours. These events were preceded by an arsenic trioxide dose-dependent down-regulation of activated STAT proteins starting at 6 hours. We hypothesized that arsenic trioxide inhibits protein tyrosine kinases (PTK), which, among others, phosphorylate and activate STATs. We therefore studied arsenic trioxide effects on Janus kinases and on three oncogenic PTKs that are known to activate STATs [FLT3, ZNF198/fibroblast growth factor receptor 1 (FGFR1), and BCR/ABL]. Arsenic trioxide reduced STAT3 activation by Janus kinases, altered phosphorylation and electrophoretic mobility of ZNF198/fibroblast growth factor receptor 1, reduced kinase protein level, and decreased STAT3 protein phosphorylation. Arsenic trioxide also reduced the phosphorylation of BCR/ABL and FLT3 with corresponding decreased STAT5 phosphorylation.
Conclusions: These results suggest a selective activity of arsenic trioxide on PTKs and will assist in developing clinical trials in AML.
Figures



References
-
- Benekli M, Baer MR, Baumann H, Wetzler M. Signal transducer and activator of transcription proteins in leukemias. Blood. 2003;101:2940–2954. - PubMed
-
- Xia Z, Baer MR, Block AW, Baumann H, Wetzler M. Expression of signal transducers and activators of transcription proteins in acute myeloid leukemia blasts. Cancer Res. 1998;58:3173–3180. - PubMed
-
- Benekli M, Xia Z, Donohue KA, Ford LA, Pixley LA, Baer MR, Baumann H, Wetzler M. Constitutive activity of signal transducer and activator of transcription 3 protein in acute myeloid leukemia blasts is associated with short disease-free survival. Blood. 2002;99:252–257. - PubMed
-
- Shen ZX, Chen GQ, Ni JH, Li XS, Xiong SM, Qiu QY, Zhu J, Tang W, Sun GL, Yang KQ, Chen Y, Zhou L, Fang ZW, Wang YT, Ma J, Zhang P, Zhang TD, Chen SJ, Chen Z, Wang ZY. Use of arsenic trioxide (As2O3) in the treatment of acute promyelocytic leukemia (APL): II. Clinical efficacy and pharmacokinetics in relapsed patients. Blood. 1997;89:3354–3360. - PubMed
-
- Bachleitner-Hofmann T, Gisslinger B, Grumbeck E, Gisslinger H. Arsenic trioxide and ascorbic acid: synergy with potential implications for the treatment of acute myeloid leukaemia? Br J Haematol. 2001;112:783–786. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
