

Spinal NK-1 receptor expressing neurons mediate opioid-induced hyperalgesia and antinociceptive tolerance via activation of descending pathways
- PMID: 17123731
- PMCID: PMC4028682
- DOI: 10.1016/j.pain.2006.09.033
Spinal NK-1 receptor expressing neurons mediate opioid-induced hyperalgesia and antinociceptive tolerance via activation of descending pathways
Abstract
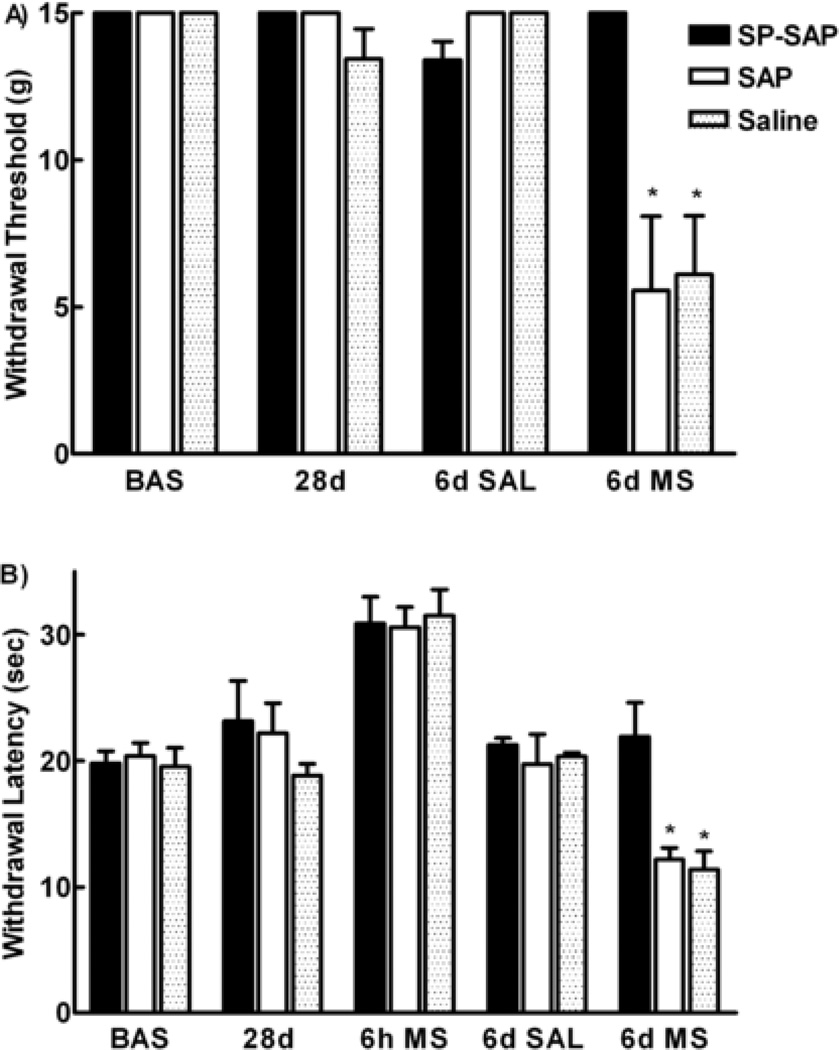
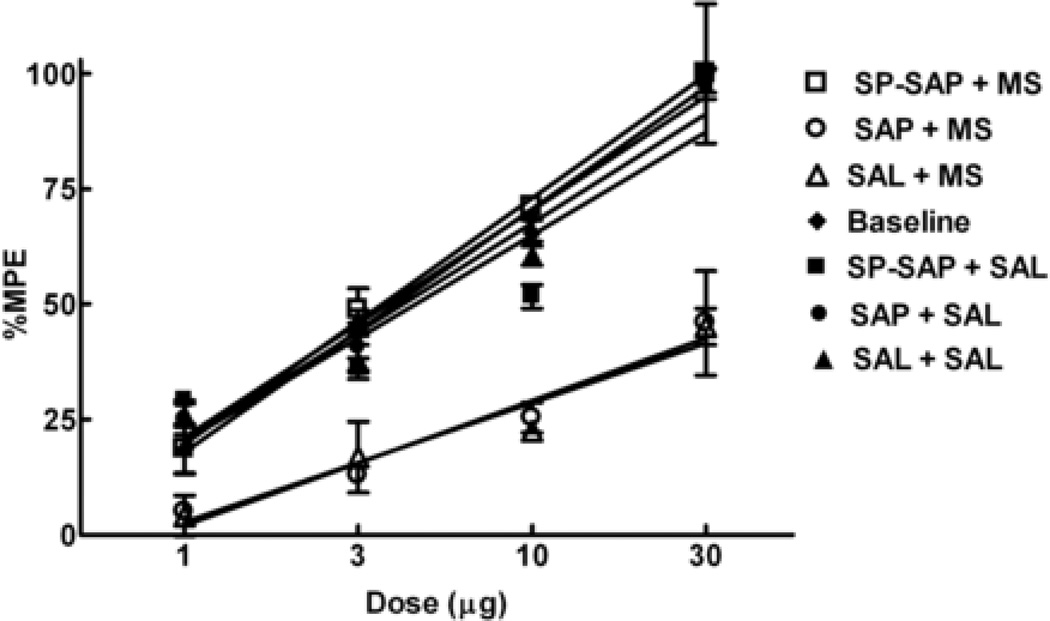
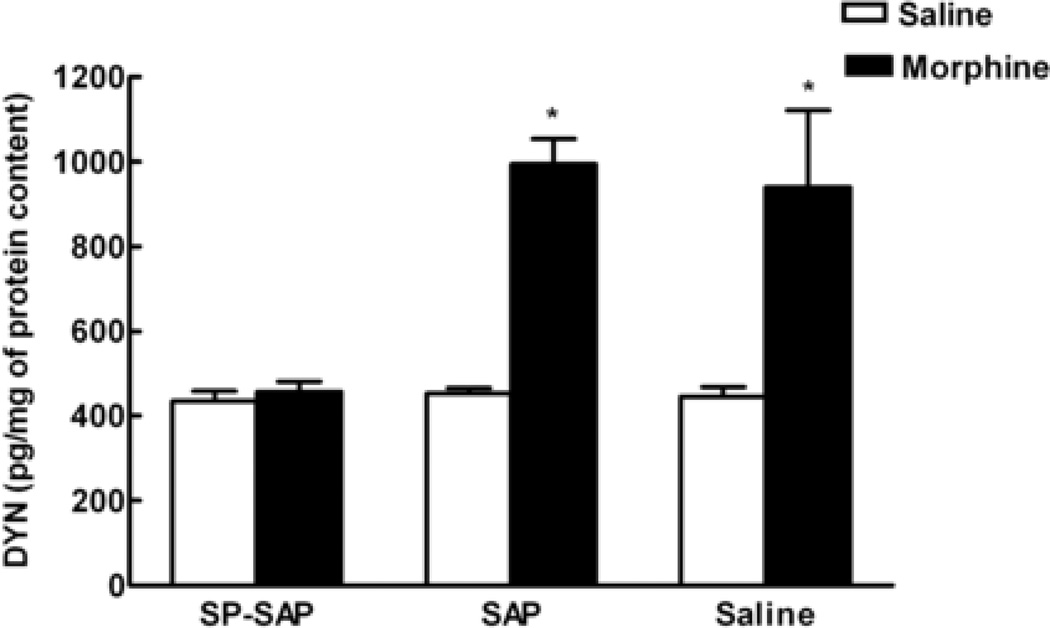
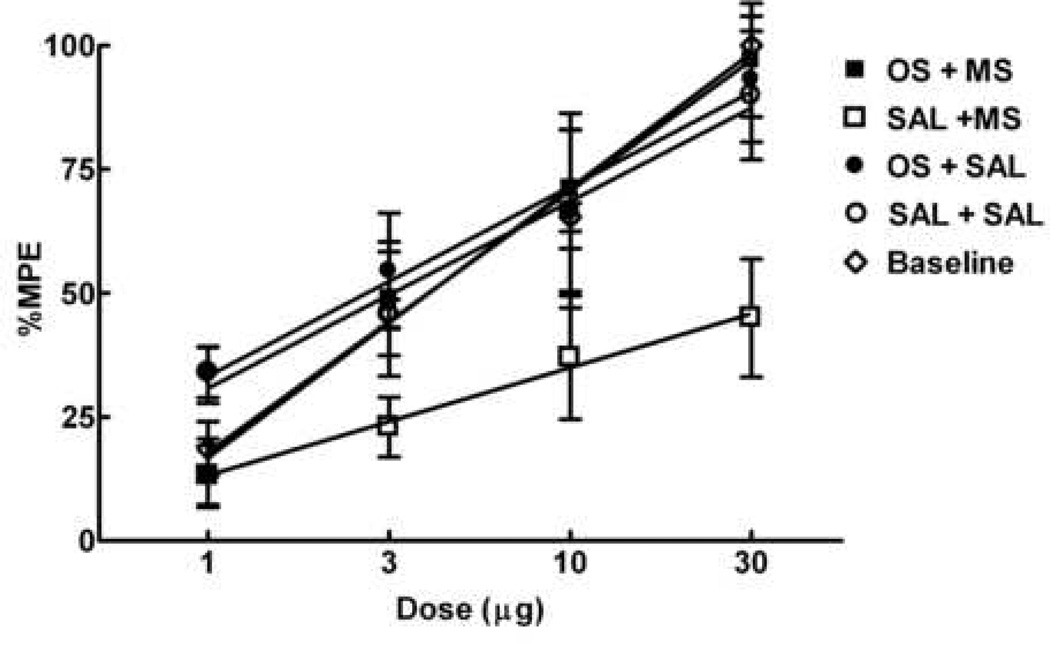
Opioids can induce hyperalgesia in humans and in animals. Mechanisms of opiate-induced hyperalgesia and possibly of spinal antinociceptive tolerance may be linked to pronociceptive adaptations occurring at multiple levels of the nervous system including activation of descending facilitatory influences from the brainstem, spinal neuroplasticity, and changes in primary afferent fibers. Here, the role of NK-1 receptor expressing cells in the spinal dorsal horn in morphine-induced hyperalgesia and spinal antinociceptive tolerance was assessed by ablating these cells with intrathecal injection of SP-saporin (SP-SAP). Ablation of NK-1 receptor expressing cells prevented (a) morphine-induced thermal and mechanical hypersensitivity, (b) increased touch-evoked spinal FOS expression, (c) upregulation of spinal dynorphin content and (d) the rightward displacement of the spinal morphine antinociceptive dose-response curve (i.e., tolerance). Morphine-induced hyperalgesia and antinociceptive tolerance were also blocked by spinal administration of ondansetron, a serotonergic receptor antagonist. Thus, NK-1 receptor expressing neurons play a critical role in sustained morphine-induced neuroplastic changes which underlie spinal excitability reflected as thermal and tactile hypersensitivity to peripheral stimuli, and to reduced antinociceptive actions of spinal morphine (i.e., antinociceptive tolerance). Ablation of these cells likely eliminates the ascending limb of a spinal-bulbospinal loop that engages descending facilitation and elicits subsequent spinal neuroplasticity. The data may provide a basis for understanding mechanisms of prolonged pain which can occur in the absence of tissue injury.
Figures


References
-
- Abbadie C, Besson JM. C-fos expression in rat lumbar spinal cord following peripheral stimulation in adjuvant arthritic and normal rats. Brain Res. 1993;607:195–204. - PubMed
-
- Ali NM. Hyperalgesic response in a patient receiving high concentrations of spinal morphine. Anesthesiology. 1986;65:449. - PubMed
-
- Angst MS, Clark JD. Opioid-induced hyperalgesia: A qualitative systematic review. Anesthesiology. 2006;3:570–587. - PubMed
-
- Arner S, Meyerson BA. Lack of analgesic effect of opioids on neuropathic and idiopathic forms of pain. Pain. 1986;33:11–23. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
