

Neurotransmitter mechanisms mediating low-glucose signalling in cocultures and fresh tissue slices of rat carotid body
- PMID: 17124268
- PMCID: PMC2151341
- DOI: 10.1113/jphysiol.2006.121871
Neurotransmitter mechanisms mediating low-glucose signalling in cocultures and fresh tissue slices of rat carotid body
Abstract
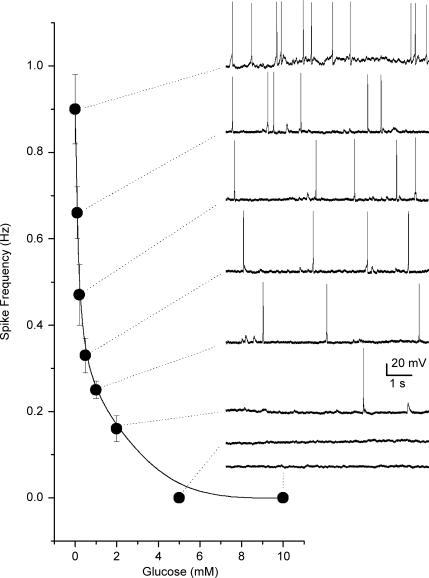
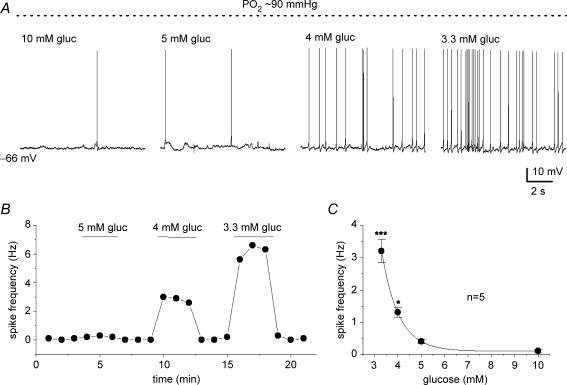
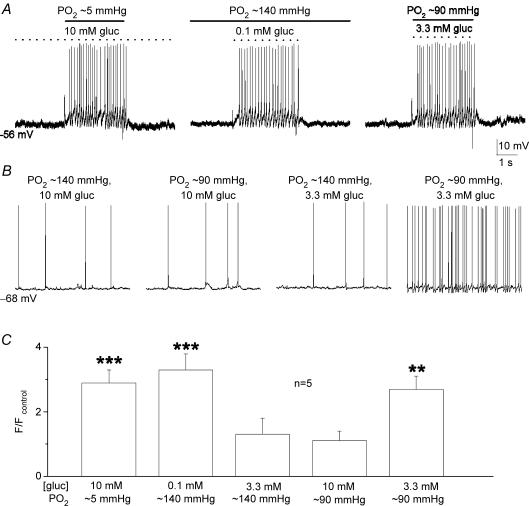
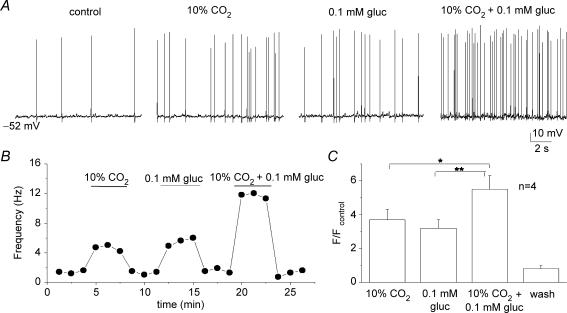
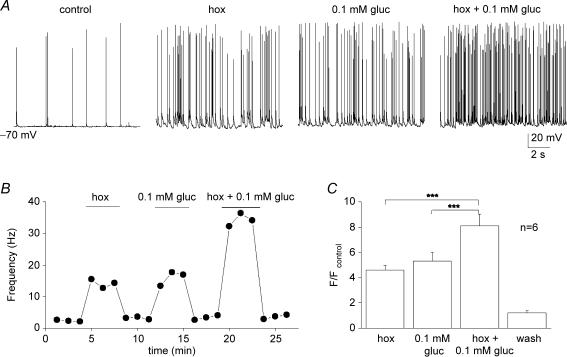
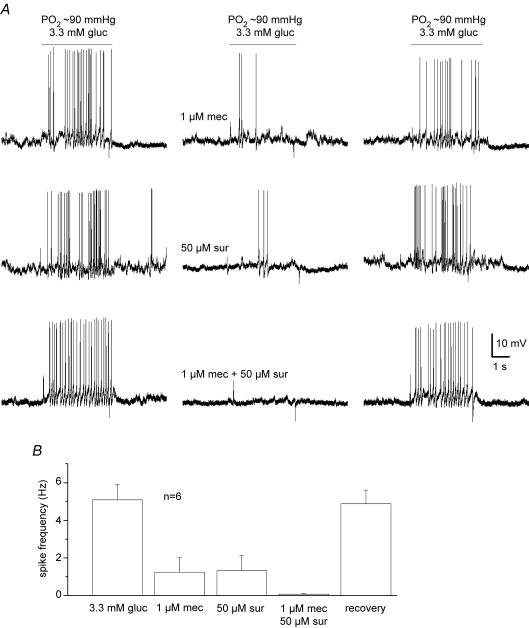
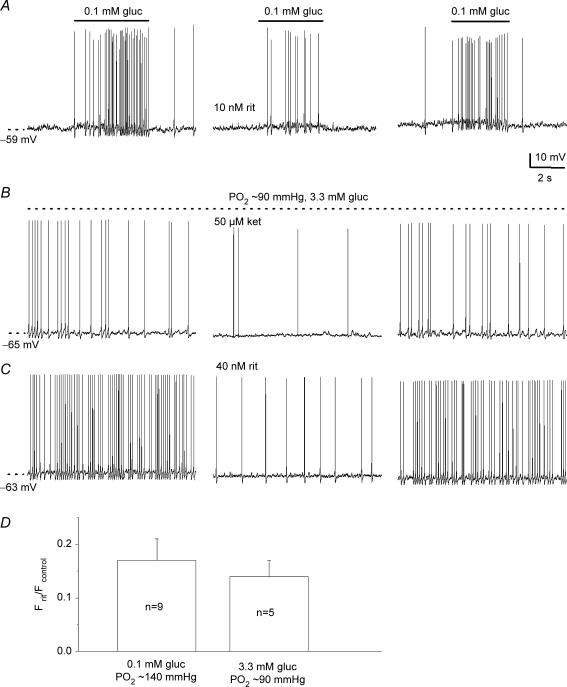
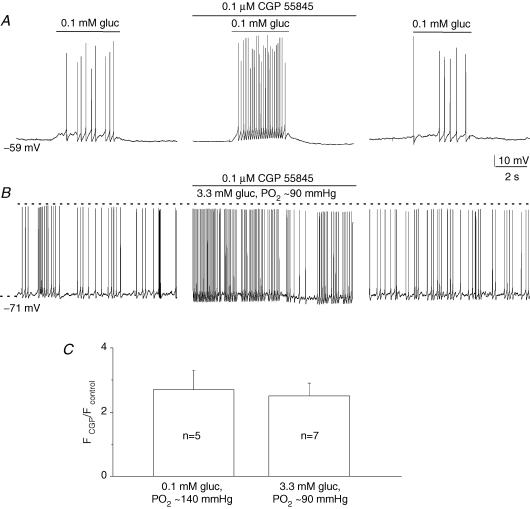
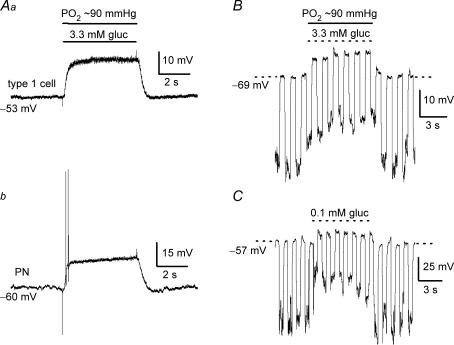
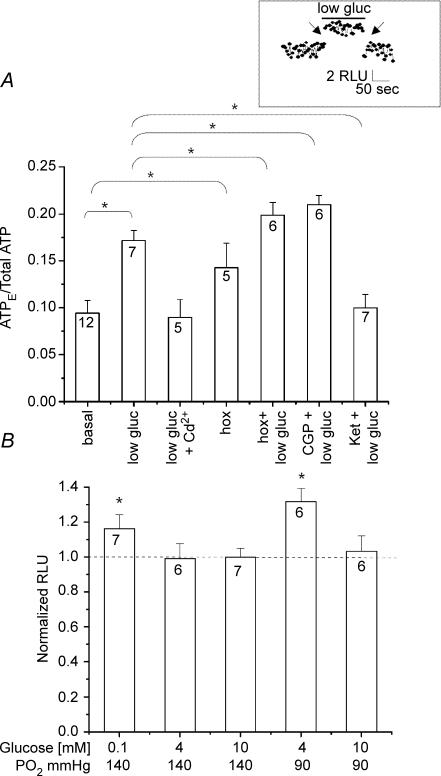
The mammalian carotid body (CB) is a polymodal chemosensor which can detect low blood glucose (hypoglycaemia), leading to increased afferent discharge and activation of counter-regulatory autonomic pathways. The underlying neurotransmitter mechanisms are unknown and controversy surrounds whether the action of low glucose is direct or indirect. To address this, we used a coculture model containing functional chemosensory units of rat CB receptor (type I) cell clusters and afferent petrosal neurones (PN). During perforated-patch, whole-cell recordings, low glucose (0-2 mM) stimulated sensory discharge in cocultured PN. When the background P(O2) was lowered to levels typical of arterial blood (approximately 90 mmHg), robust PN chemoexcitation could be induced by physiological hypoglycaemia (3.3-4 mM glucose). These sensory responses were reversibly inhibited by a combination of purinergic (suramin, 50 microM) and nicotinic (mecamylamine, 1 microM) receptor blockers, suggesting that transmission depended on corelease of ATP and ACh. Hypoglycaemic responses were additive with those evoked by hypoxia or hypercapnia; further, they could be potentiated by the GABAB receptor blocker (CGP 55845) and inhibited by 5-HT2A receptor blockers (ketanserin or ritanserin). During paired simultaneous recordings from a PN and a type I cell in an adjacent cluster, the afferent PN response coincided with type I cell depolarization, which was associated with a decrease in input resistance. In fresh tissue slices of rat CB, low glucose stimulated ATP secretion as determined by the luciferin-luciferase assay; this secretion was cadmium sensitive, potentiated by CGP 55845, and inhibited by ketanserin. Taken together these data indicate that CB receptors act as direct glucosensors, and that processing of hypoglycaemia utilizes similar neurotransmitter and neuromodulatory mechanisms as hypoxia.
Figures
Comment in
-
How sweet it is: sensing low glucose in the carotid body.J Physiol. 2007 Feb 1;578(Pt 3):627. doi: 10.1113/jphysiol.2006.126250. Epub 2006 Dec 14. J Physiol. 2007. PMID: 17170041 Free PMC article. No abstract available.
Similar articles
-
CO2/pH chemosensory signaling in co-cultures of rat carotid body receptors and petrosal neurons: role of ATP and ACh.J Neurophysiol. 2004 Dec;92(6):3433-45. doi: 10.1152/jn.01099.2003. Epub 2004 Mar 31. J Neurophysiol. 2004. PMID: 15056681
-
Acetylcholine contributes to hypoxic chemotransmission in co-cultures of rat type 1 cells and petrosal neurons.Respir Physiol. 1999 Apr 1;115(2):189-99. doi: 10.1016/s0034-5687(99)00017-1. Respir Physiol. 1999. PMID: 10385033
-
Postsynaptic action of GABA in modulating sensory transmission in co-cultures of rat carotid body via GABA(A) receptors.J Physiol. 2009 Jan 15;587(2):329-44. doi: 10.1113/jphysiol.2008.165035. Epub 2008 Nov 24. J Physiol. 2009. PMID: 19029183 Free PMC article.
-
Neurotransmitter and neuromodulatory mechanisms at peripheral arterial chemoreceptors.Exp Physiol. 2010 Jun;95(6):657-67. doi: 10.1113/expphysiol.2009.049312. Epub 2010 Apr 1. Exp Physiol. 2010. PMID: 20360424 Review.
-
Neurotransmission and neuromodulation in the chemosensory carotid body.Auton Neurosci. 2005 Jun 15;120(1-2):1-9. doi: 10.1016/j.autneu.2005.04.008. Auton Neurosci. 2005. PMID: 15955746 Review.
Cited by
-
Carotid body chemoreceptors: physiology, pathology, and implications for health and disease.Physiol Rev. 2021 Jul 1;101(3):1177-1235. doi: 10.1152/physrev.00039.2019. Epub 2021 Feb 11. Physiol Rev. 2021. PMID: 33570461 Free PMC article. Review.
-
Cellular properties and chemosensory responses of the human carotid body.J Physiol. 2013 Dec 15;591(24):6157-73. doi: 10.1113/jphysiol.2013.263657. Epub 2013 Oct 28. J Physiol. 2013. PMID: 24167224 Free PMC article.
-
The effects of hypo- and hyperglycaemia on the hypoxic ventilatory response in humans.J Physiol. 2007 Jul 15;582(Pt 2):859-69. doi: 10.1113/jphysiol.2007.130112. Epub 2007 May 3. J Physiol. 2007. PMID: 17478538 Free PMC article.
-
Is the Carotid Body a Metabolic Monitor?Adv Exp Med Biol. 2015;860:153-9. doi: 10.1007/978-3-319-18440-1_17. Adv Exp Med Biol. 2015. PMID: 26303477 Free PMC article.
-
Carotid body: an emerging target for cardiometabolic co-morbidities.Exp Physiol. 2023 May;108(5):661-671. doi: 10.1113/EP090090. Epub 2023 Mar 30. Exp Physiol. 2023. PMID: 36999224 Free PMC article. Review.
References
-
- Alvarez-Buylla R, de Alvarez-Buylla ER. Carotid sinus receptors participate in glucose homeostasis. Respir Physiol. 1988;72:347–360. - PubMed
-
- Buniel MC, Schilling WP, Kunze DL. Distribution of transient receptor potential channels in the rat carotid chemosensory pathway. J Comp Neurol. 2003;464:404–413. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical