

Structural basis for ribosome recruitment and manipulation by a viral IRES RNA
- PMID: 17124290
- PMCID: PMC2669756
- DOI: 10.1126/science.1133281
Structural basis for ribosome recruitment and manipulation by a viral IRES RNA
Abstract
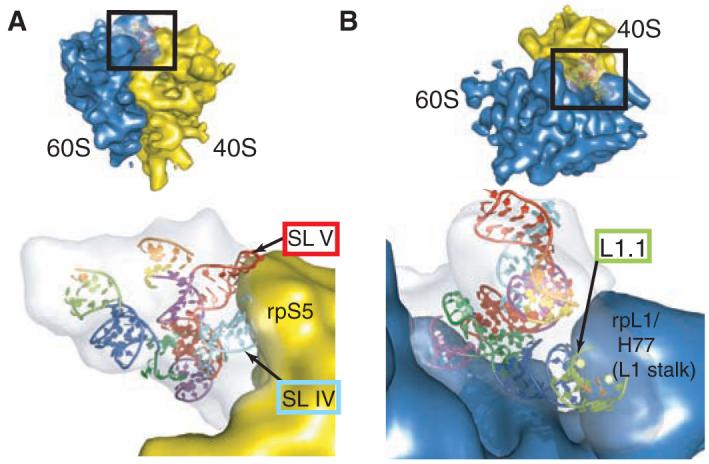
Canonical cap-dependent translation initiation requires a large number of protein factors that act in a stepwise assembly process. In contrast, internal ribosomal entry sites (IRESs) are cis-acting RNAs that in some cases completely supplant these factors by recruiting and activating the ribosome using a single structured RNA. Here we present the crystal structures of the ribosome-binding domain from a Dicistroviridae intergenic region IRES at 3.1 angstrom resolution, providing a view of the prefolded architecture of an all-RNA translation initiation apparatus. Docking of the structure into cryo-electron microscopy reconstructions of an IRES-ribosome complex suggests a model for ribosome manipulation by a dynamic IRES RNA.
Figures



References
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
