

Specific effects of potassium ion binding on wild-type and L358P cytochrome P450cam
- PMID: 17128977
- PMCID: PMC1764623
- DOI: 10.1021/bi0617355
Specific effects of potassium ion binding on wild-type and L358P cytochrome P450cam
Abstract
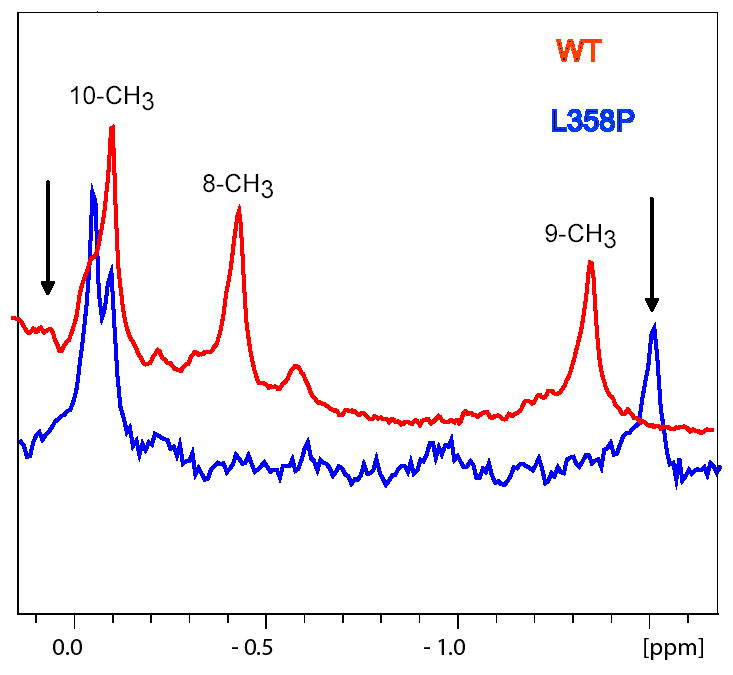
The camphor monoxygenase cytochrome P450cam (CYP101) requires potassium ion (K+) to drive formation of the characteristic high-spin state of the heme Fe+3 upon substrate binding. Amide 1H, 15N correlations in perdeuterated [U-15N] CYP101 were monitored as a function of K+ concentration by 2D-TROSY-HSQC in both camphor-bound oxidized (CYP-S) and camphor- and CO-bound reduced CYP101 (CYP-S-CO). In both forms, K+-induced spectral perturbations are detected in the vicinity of the K+ binding site proposed from crystallographic structures, but are larger and more widespread structurally in CYP-S than in CYP-S-CO. In CYP-S-CO, K+-induced perturbations occur primarily near the proposed K+ binding site in the B-B' loop and B' helix, which are also perturbed by binding of effector, putidaredoxin (Pdx). The spectral effects of K+ binding in CYP-S-CO oppose those observed upon Pdxr titration. However, Pdxr titration of CYP-S-CO in the absence of K+ results in multiple conformations. The spin-state equilibrium in the L358P mutant of CYP101 is more sensitive to K+ concentration than WT CYP101, consistent with a hypothesis that L358P preferentially populates conformations enforced by Pdx binding in WT CYP101. Thallium(I), a K+ mimic, minimizes the effects of Pdx titration on the NMR spectrum of CYP-S-CO, but is competent to replace K+ in driving the formation of high-spin CYP-S. These observations suggest that the role of K+ is to stabilize conformers of CYP-S that drive the spin-state change prior to the first electron transfer, and that K+ stabilizes the CYP-S-CO conformer that interacts with Pdx. However, upon binding of Pdx, further conformational changes occur that disfavor K+ binding.
Figures





References
-
- Britt BM. For enzymes, bigger is better. Biophys Chem. 1997;69:63–70. - PubMed
-
- Mueller EJ, Loida PJ, Sligar SG. Cytochrome P450: Structure, Function and Biochemistry. 1995. pp. 83–124.
-
- Lange R, Bonfils C, Debey P. The low-spin reversible highspin transition equilibrium of camphor-bound cytochrome P-450. Effects of medium and temperature on equilibrium data. Eur J Biochem. 1977;79:623–628. - PubMed
-
- Lange R, Pierre J, Debey P. Visible and ultraviolet spectral transitions of camphor-bound cytochrome P-450. A comprehensive study. Eur J Biochem. 1980;107:441–445. - PubMed
-
- Hui Bon Hoa G, Marden MC. The pressure dependence of the spin equilibrium of camphor-bound cytochrome P450. Eur J Biochem. 1982;124:311–315. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
