

A deep-branching clade of retrovirus-like retrotransposons in bdelloid rotifers
- PMID: 17129685
- PMCID: PMC1839950
- DOI: 10.1016/j.gene.2006.09.025
A deep-branching clade of retrovirus-like retrotransposons in bdelloid rotifers
Abstract
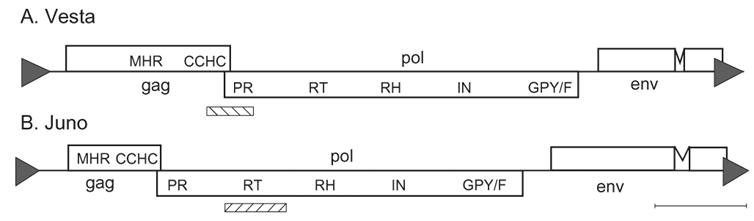
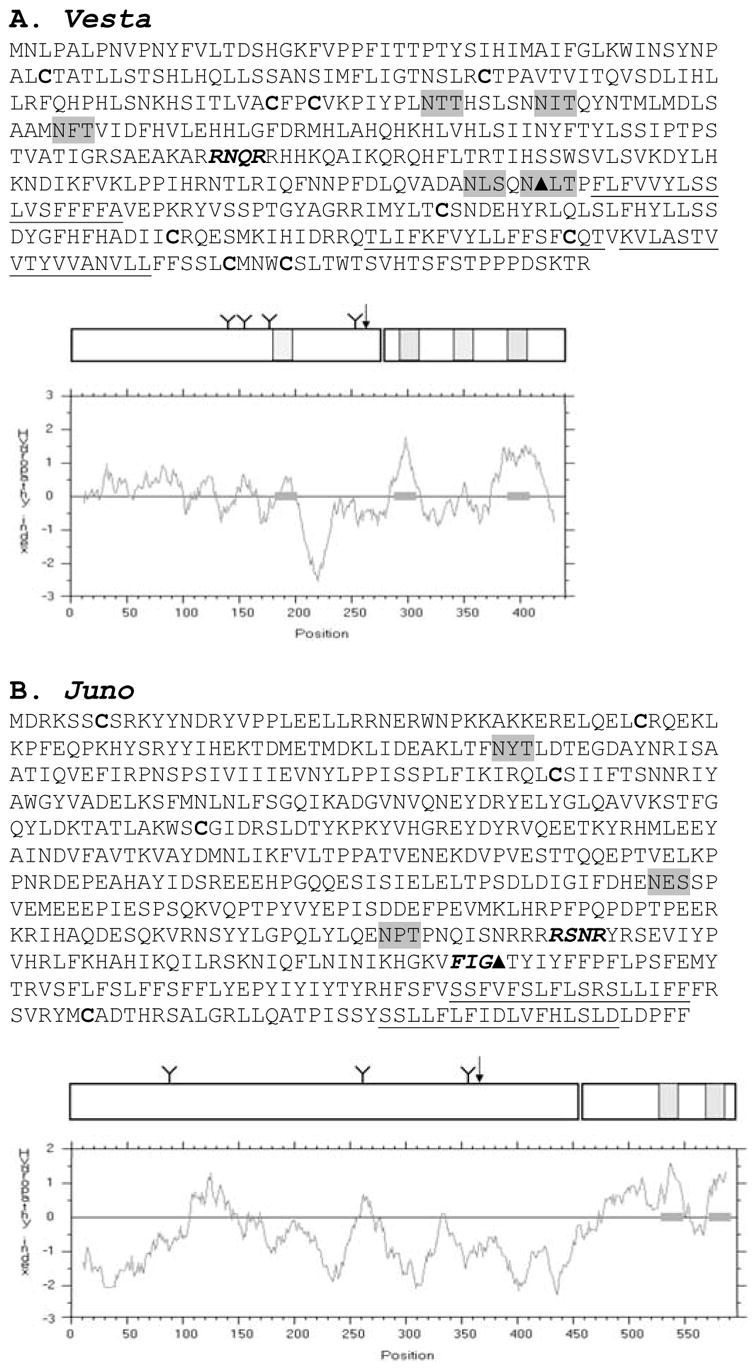
Rotifers of class Bdelloidea, a group of aquatic invertebrates in which males and meiosis have never been documented, are also unusual in their lack of multicopy LINE-like and gypsy-like retrotransposons, groups inhabiting the genomes of nearly all other metazoans. Bdelloids do contain numerous DNA transposons, both intact and decayed, and domesticated Penelope-like retroelements Athena, concentrated at telomeric regions. Here we describe two LTR retrotransposons, each found at low copy number in a different bdelloid species, which define a clade different from previously known clades of LTR retrotransposons. Like bdelloid DNA transposons and Athena, these elements are found preferentially in telomeric regions. Unlike bdelloid DNA transposons, many of which are decayed, the newly described elements, named Vesta and Juno, inhabiting the genomes of Philodina roseola and Adineta vaga, respectively, appear to be intact and represent recent insertions, possibly from an exogenous source. We describe the retrovirus-like structure of the new elements, containing gag, pol, and env-like open reading frames, and discuss their possible origins, transmission, and behavior in bdelloid genomes.
Figures



References
-
- Arkhipova IR, Mazo AM, Cherkasova VA, Gorelova TV, Schuppe NG, Ilyin YV. The steps of reverse transcription of Drosophila mobile dispersed genetic elements and U3-R-U5 structure of their LTRs. Cell. 1986;44:555–563. - PubMed
-
- Arkhipova IR, Lyubomirskaya NV, Ilyin YV. Drosophila retrotransposons. Austin, TX: RG Landes Co; 1995.
-
- Arkhipova I, Meselson M. Deleterious transposable elements and the extinction of asexuals. Bioessays. 2005a;27:76–85. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
