

Inactivation of CUG-BP1/CELF1 causes growth, viability, and spermatogenesis defects in mice
- PMID: 17130239
- PMCID: PMC1800704
- DOI: 10.1128/MCB.01009-06
Inactivation of CUG-BP1/CELF1 causes growth, viability, and spermatogenesis defects in mice
Abstract
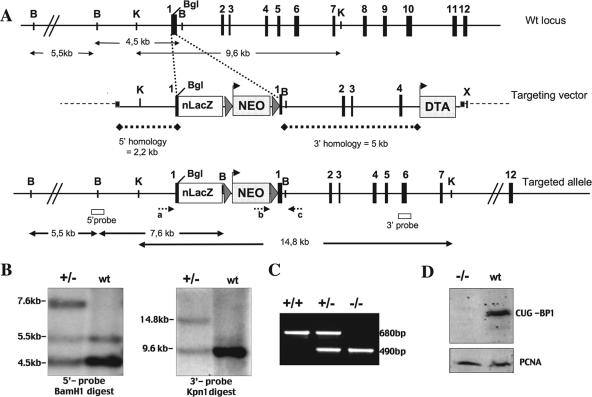
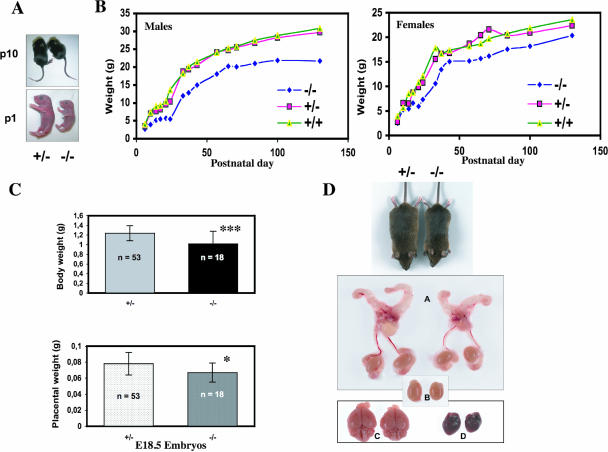
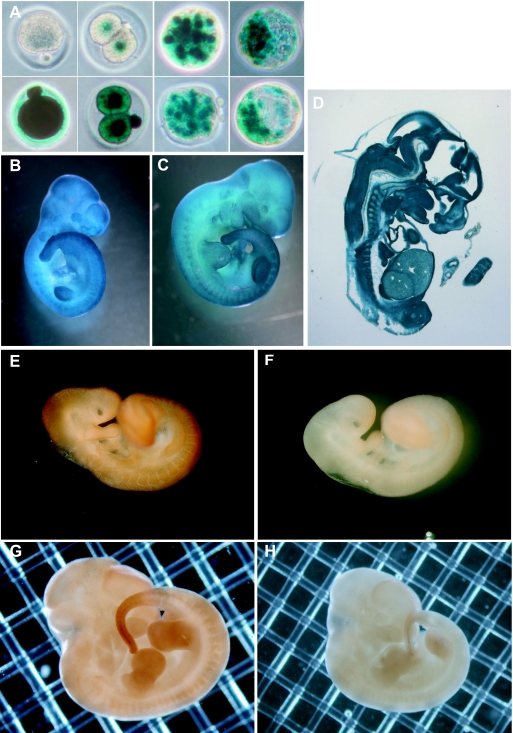
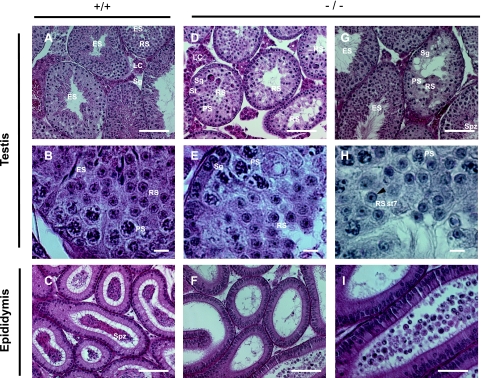
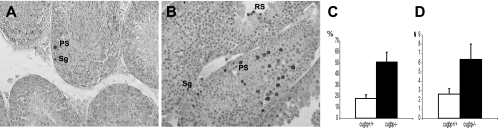
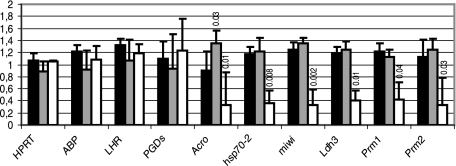
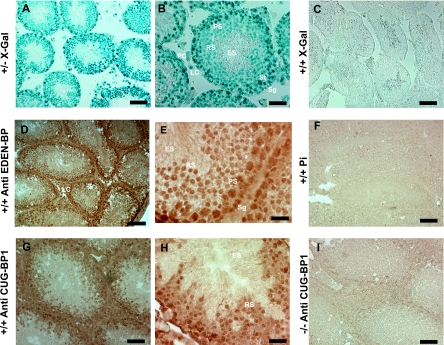
CUG-BP1/CELF1 is a multifunctional RNA-binding protein involved in the regulation of alternative splicing and translation. To elucidate its role in mammalian development, we produced mice in which the Cugbp1 gene was inactivated by homologous recombination. These Cugbp1(-/-) mice were viable, although a significant portion of them did not survive after the first few days of life. They displayed growth retardation, and most Cugbp1(-/-) males and females exhibited impaired fertility. Male infertility was more thoroughly investigated. Histological examination of testes from Cugbp1(-/-) males showed an arrest of spermatogenesis that occurred at step 7 of spermiogenesis, before spermatid elongation begins, and an increased apoptosis. A quantitative reverse transcriptase PCR analysis showed a decrease of all the germ cell markers tested but not of Sertoli and Leydig markers, suggesting a general decrease in germ cell number. In wild-type testes, CUG-BP1 is expressed in germ cells from spermatogonia to round spermatids and also in Sertoli and Leydig cells. These findings demonstrate that CUG-BP1 is required for completion of spermatogenesis.
Figures
References
-
- Barreau, C., L. Paillard, A. Mereau, and H. B. Osborne. 2006. Mammalian CELF/Bruno-like RNA-binding proteins: molecular characteristics and biological functions. Biochimie 88:515-525. - PubMed
-
- Barreau, C., T. Watrin, H. Beverley Osborne, and L. Paillard. 2006. Protein expression is increased by a class III AU-rich element and tethered CUG-BP1. Biochem. Biophys. Res. Commun. 347:723-730. - PubMed
-
- Behr, R., and G. F. Weinbauer. 2001. cAMP response element modulator (CREM): an essential factor for spermatogenesis in primates? Int. J. Androl. 24:126-135. - PubMed
-
- Blendy, J. A., K. H. Kaestner, G. F. Weinbauer, E. Nieschlag, and G. Schutz. 1996. Severe impairment of spermatogenesis in mice lacking the CREM gene. Nature 380:162-165. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases