

A driving and coupling "Pac-Man" mechanism for chromosome poleward translocation in anaphase A
- PMID: 17130449
- PMCID: PMC1693682
- DOI: 10.1073/pnas.0608962103
A driving and coupling "Pac-Man" mechanism for chromosome poleward translocation in anaphase A
Abstract
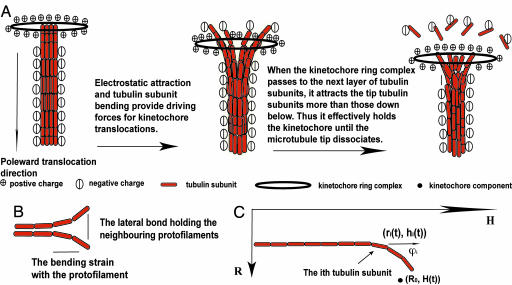
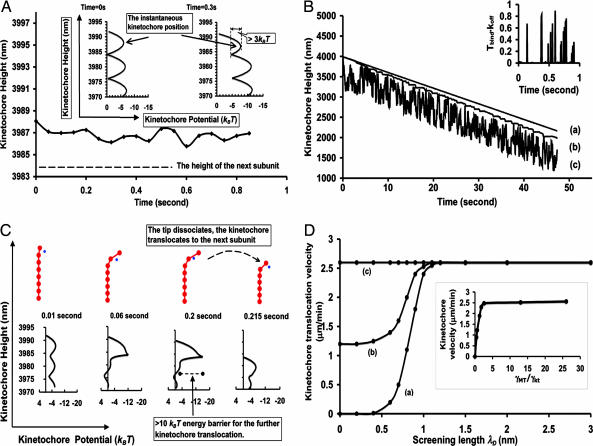
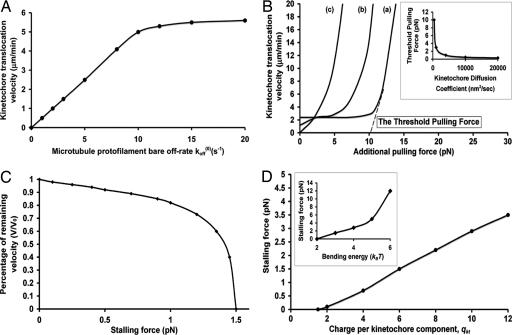
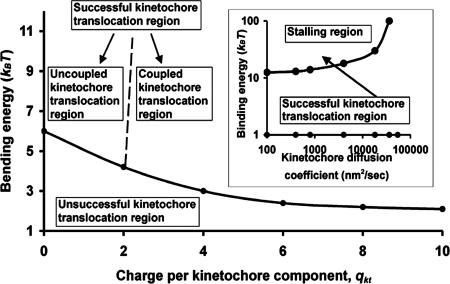
During mitosis, chromatid harnesses its kinetochore translocation at the depolymerizing microtubule ends for its poleward movement in anaphase A. The force generation mechanism for such movement remains unknown. Analysis of the current experimental results shows that the bending energy release from the bound tubulin subunits alone cannot provide sufficient driving force. Additional contribution from effective electrostatic attractions between the kinetochore and the microtubule is needed for kinetochore translocation. Interestingly, as the kinetochore moves to inside the microtubule, the microtubule tip is free to bend outward so that the instantaneous distance between the kinetochore and the microtubule tip is much closer than the rest of the microtubule. This close contact yields much larger electrostatic attraction than that from the rest of the microtubule under physiological ionic conditions. As a result, the effective electrostatic interaction hinders the further kinetochore poleward translocation until the microtubule tip dissociates. Thus, the kinetochore translocation is strongly coupled at the depolymerizing microtubule end. This driving-coupling mechanism indicates that the kinetochore velocity is largely controlled by the microtubule dissociation rate, which explains the insensitivity of kinetochore velocity to its viscous drag and the large redundancy in its stalling force.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
Pac-Man does not resolve the enduring problem of anaphase chromosome movement.Protoplasma. 2001;215(1-4):16-20. doi: 10.1007/BF01280300. Protoplasma. 2001. PMID: 11732055 Review.
-
Direct observation of microtubule dynamics at kinetochores in Xenopus extract spindles: implications for spindle mechanics.J Cell Biol. 2003 Aug 4;162(3):377-82. doi: 10.1083/jcb.200301088. J Cell Biol. 2003. PMID: 12900391 Free PMC article.
-
A mechanomolecular model for the movement of chromosomes during mitosis driven by a minimal kinetochore bicyclic cascade.J Theor Biol. 2010 Apr 21;263(4):455-70. doi: 10.1016/j.jtbi.2009.12.023. Epub 2010 Jan 4. J Theor Biol. 2010. PMID: 20043924
-
Direct visualization of microtubule flux during metaphase and anaphase in crane-fly spermatocytes.Mol Biol Cell. 2004 Dec;15(12):5724-32. doi: 10.1091/mbc.e04-08-0750. Epub 2004 Oct 6. Mol Biol Cell. 2004. PMID: 15469981 Free PMC article.
-
Molecular architecture of the kinetochore-microtubule interface.Nat Rev Mol Cell Biol. 2008 Jan;9(1):33-46. doi: 10.1038/nrm2310. Nat Rev Mol Cell Biol. 2008. PMID: 18097444 Review.
Cited by
-
The Dam1 ring binds microtubules strongly enough to be a processive as well as energy-efficient coupler for chromosome motion.Proc Natl Acad Sci U S A. 2008 Oct 7;105(40):15423-8. doi: 10.1073/pnas.0807859105. Epub 2008 Sep 29. Proc Natl Acad Sci U S A. 2008. PMID: 18824692 Free PMC article.
-
A mechanobiochemical mechanism for monooriented chromosome oscillation in mitosis.Proc Natl Acad Sci U S A. 2007 Oct 9;104(41):16104-9. doi: 10.1073/pnas.0707689104. Epub 2007 Oct 2. Proc Natl Acad Sci U S A. 2007. PMID: 17911248 Free PMC article.
-
Highly Transient Molecular Interactions Underlie the Stability of Kinetochore-Microtubule Attachment During Cell Division.Cell Mol Bioeng. 2013 Dec 13;6(4):10.1007/s12195-013-0309-4. doi: 10.1007/s12195-013-0309-4. Cell Mol Bioeng. 2013. PMID: 24376473 Free PMC article.
-
An integrated mechanobiochemical feedback mechanism describes chromosome motility from prometaphase to anaphase in mitosis.Proc Natl Acad Sci U S A. 2008 Sep 16;105(37):13752-7. doi: 10.1073/pnas.0807007105. Epub 2008 Sep 9. Proc Natl Acad Sci U S A. 2008. PMID: 18780795 Free PMC article.
-
Electrostatic forces drive poleward chromosome motions at kinetochores.Cell Div. 2016 Oct 28;11:14. doi: 10.1186/s13008-016-0026-1. eCollection 2016. Cell Div. 2016. PMID: 27807465 Free PMC article.
References
-
- Maiato H, Deluca J, Salmon ED, Earnshaw WC. J Cell Sci. 2004;117:5461–5477. - PubMed
-
- Desai A, Mitchison TJ. Annu Rev Cell Dev Biol. 1997;13:83–117. - PubMed
-
- McIntosh JR, Grishchuk EL, West RR. Annu Rev Cell Dev Biol. 2002;18:193–219. - PubMed
-
- Westermann S, Avila-Sakar A, Wang HW, Niederstrasser H, Drubin DG, Nogales E, Barnes G. Mol Cell. 2005;17:277–290. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
