

An S6 mutation in BK channels reveals beta1 subunit effects on intrinsic and voltage-dependent gating
- PMID: 17130522
- PMCID: PMC2151602
- DOI: 10.1085/jgp.200609596
An S6 mutation in BK channels reveals beta1 subunit effects on intrinsic and voltage-dependent gating
Abstract
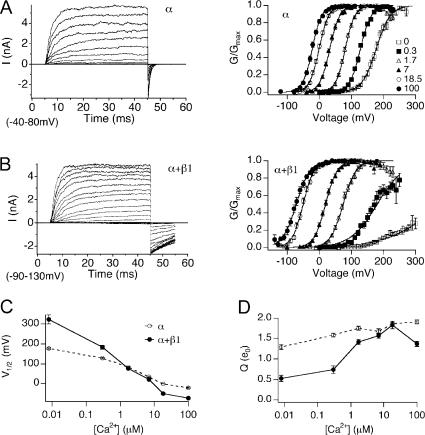
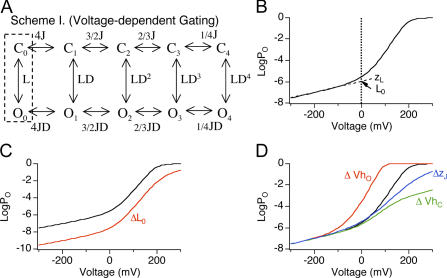
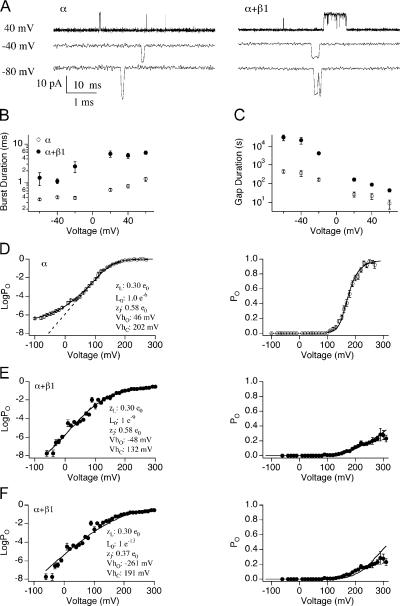
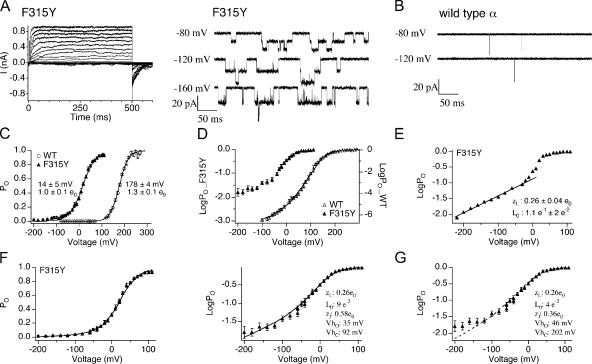
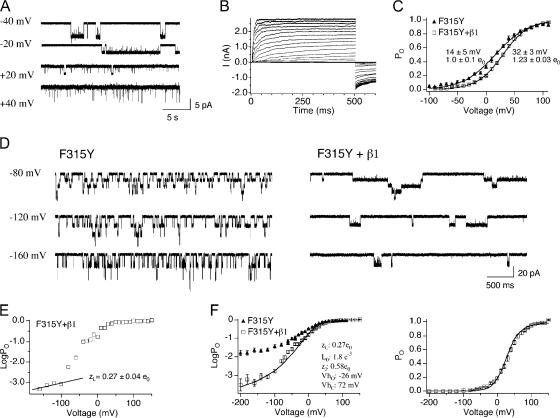
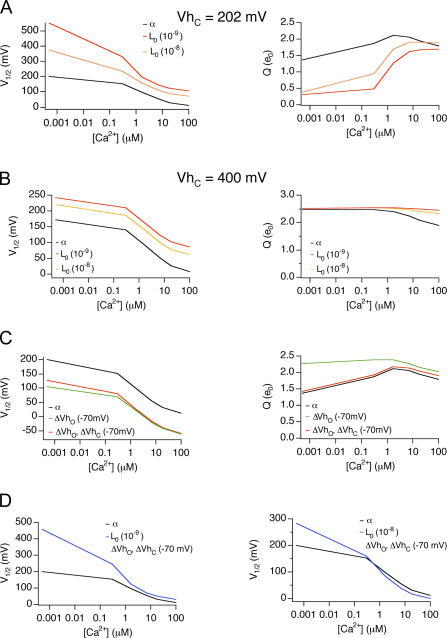
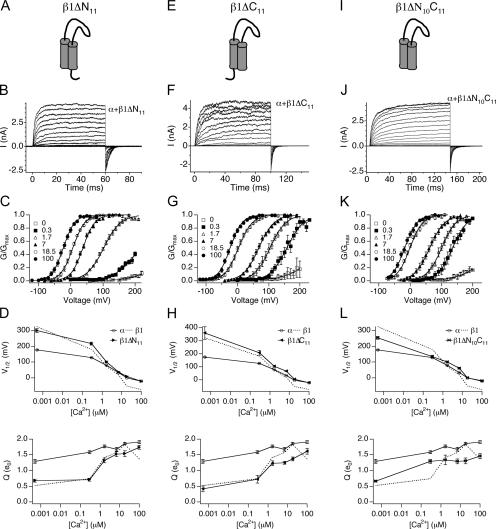
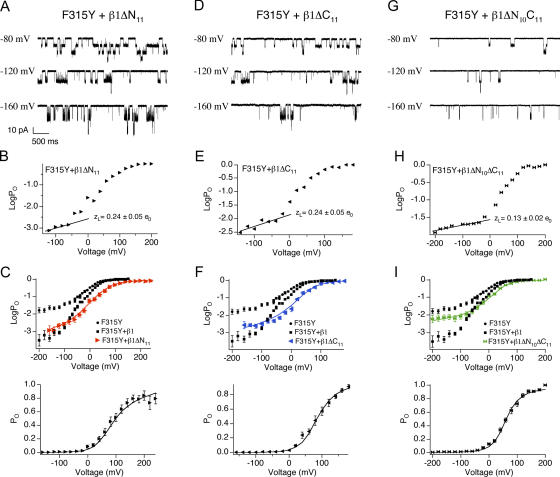
Large conductance, Ca(2+)- and voltage-activated K(+) (BK) channels are exquisitely regulated to suit their diverse roles in a large variety of physiological processes. BK channels are composed of pore-forming alpha subunits and a family of tissue-specific accessory beta subunits. The smooth muscle-specific beta1 subunit has an essential role in regulating smooth muscle contraction and modulates BK channel steady-state open probability and gating kinetics. Effects of beta1 on channel's gating energetics are not completely understood. One of the difficulties is that it has not yet been possible to measure the effects of beta1 on channel's intrinsic closed-to-open transition (in the absence of voltage sensor activation and Ca(2+) binding) due to the very low open probability in the presence of beta1. In this study, we used a mutation of the alpha subunit (F315Y) that increases channel openings by greater than four orders of magnitude to directly compare channels' intrinsic open probabilities in the presence and absence of the beta1 subunit. Effects of beta1 on steady-state open probabilities of both wild-type alpha and the F315Y mutation were analyzed using the dual allosteric HA model. We found that mouse beta1 has two major effects on channel's gating energetics. beta1 reduces the intrinsic closed-to-open equilibrium that underlies the inhibition of BK channel opening seen in submicromolar Ca(2+). Further, P(O) measurements at limiting slope allow us to infer that beta1 shifts open channel voltage sensor activation to negative membrane potentials, which contributes to enhanced channel opening seen at micromolar Ca(2+) concentrations. Using the F315Y alpha subunit with deletion mutants of beta1, we also demonstrate that the small N- and C-terminal intracellular domains of beta1 play important roles in altering channel's intrinsic opening and voltage sensor activation. In summary, these results demonstrate that beta1 has distinct effects on BK channel intrinsic gating and voltage sensor activation that can be functionally uncoupled by mutations in the intracellular domains.
Figures
References
-
- Brenner, R., T.J. Jegla, A. Wickenden, Y. Liu, and R.W. Aldrich. 2000. a. Cloning and functional characterization of novel large conductance calcium-activated potassium channel β subunits, hKCNMB3 and hKCNMB4. J. Biol. Chem. 275:6453–6461. - PubMed
-
- Brenner, R., G.J. Perez, A.D. Bonev, D.M. Eckman, J.C. Kosek, S.W. Wiler, A.J. Patterson, M.T. Nelson, and R.W. Aldrich. 2000. b. Vasoregulation by the β1 subunit of the calcium-activated potassium channel. Nature. 407:870–876. - PubMed
-
- Brenner, R., Q.H. Chen, A. Vilaythong, G.M. Toney, J.L. Noebels, and R.W. Aldrich. 2005. BK channel β4 subunit reduces dentate gyrus excitability and protects against temporal lobe seizures. Nat. Neurosci. 8:1752–1759. - PubMed
-
- Butler, A., S. Tsunoda, D.P. McCobb, A. Wei, and L. Salkoff. 1993. mSlo, a complex mouse gene encoding “maxi” calcium-activated potassium channels. Science. 261:221–224. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
