

Recurrent insertion and duplication generate networks of transposable element sequences in the Drosophila melanogaster genome
- PMID: 17134480
- PMCID: PMC1794594
- DOI: 10.1186/gb-2006-7-11-r112
Recurrent insertion and duplication generate networks of transposable element sequences in the Drosophila melanogaster genome
Abstract
Background: The recent availability of genome sequences has provided unparalleled insights into the broad-scale patterns of transposable element (TE) sequences in eukaryotic genomes. Nevertheless, the difficulties that TEs pose for genome assembly and annotation have prevented detailed, quantitative inferences about the contribution of TEs to genomes sequences.
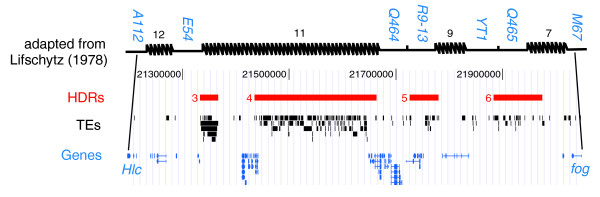
Results: Using a high-resolution annotation of TEs in Release 4 genome sequence, we revise estimates of TE abundance in Drosophila melanogaster. We show that TEs are non-randomly distributed within regions of high and low TE abundance, and that pericentromeric regions with high TE abundance are mosaics of distinct regions of extreme and normal TE density. Comparative analysis revealed that this punctate pattern evolves jointly by transposition and duplication, but not by inversion of TE-rich regions from unsequenced heterochromatin. Analysis of genome-wide patterns of TE nesting revealed a 'nesting network' that includes virtually all of the known TE families in the genome. Numerous directed cycles exist among TE families in the nesting network, implying concurrent or overlapping periods of transpositional activity.
Conclusion: Rapid restructuring of the genomic landscape by transposition and duplication has recently added hundreds of kilobases of TE sequence to pericentromeric regions in D. melanogaster. These events create ragged transitions between unique and repetitive sequences in the zone between euchromatic and beta-heterochromatic regions. Complex relationships of TE nesting in beta-heterochromatic regions raise the possibility of a co-suppression network that may act as a global surveillance system against the majority of TE families in D. melanogaster.
Figures







References
-
- Kaminker JS, Bergman CM, Kronmiller B, Carlson J, Svirskas R, Patel S, Frise E, Wheeler DA, Lewis SE, Rubin GM, et al. The transposable elements of the Drosophila melanogaster euchromatin: a genomics perspective. Genome Biol. 2002;3:RESEARCH0084. doi: 10.1186/gb-2002-3-12-research0084. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
