

Regulation of hepadnavirus reverse transcription by dynamic nucleocapsid phosphorylation
- PMID: 17135319
- PMCID: PMC1797568
- DOI: 10.1128/JVI.01671-06
Regulation of hepadnavirus reverse transcription by dynamic nucleocapsid phosphorylation
Abstract
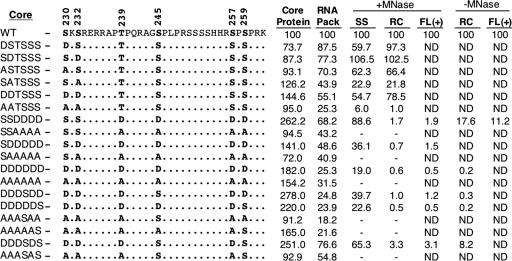
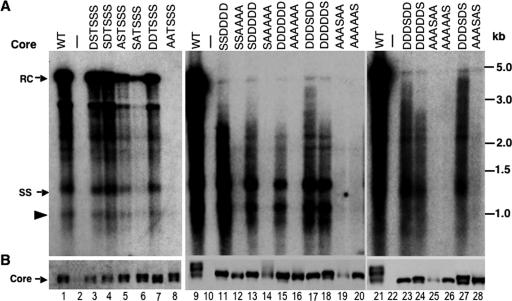
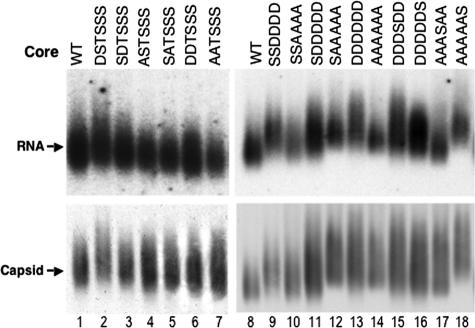
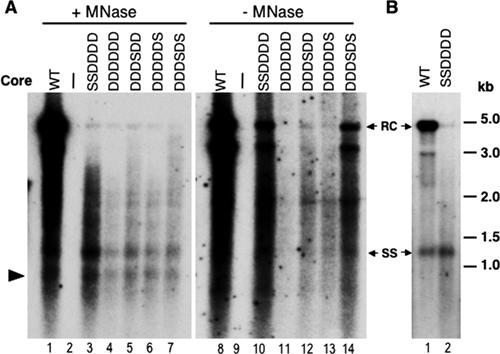
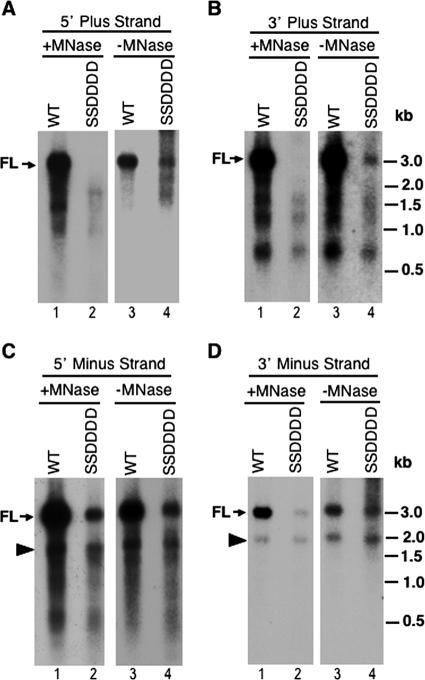
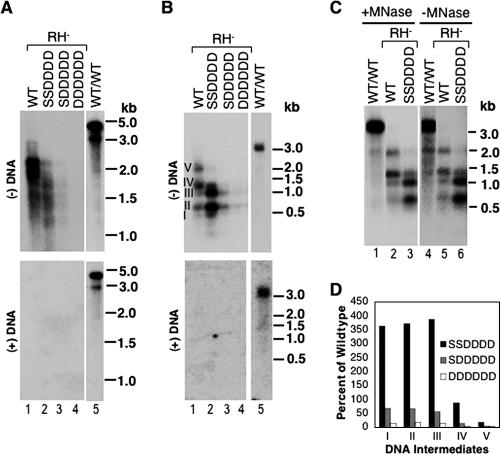
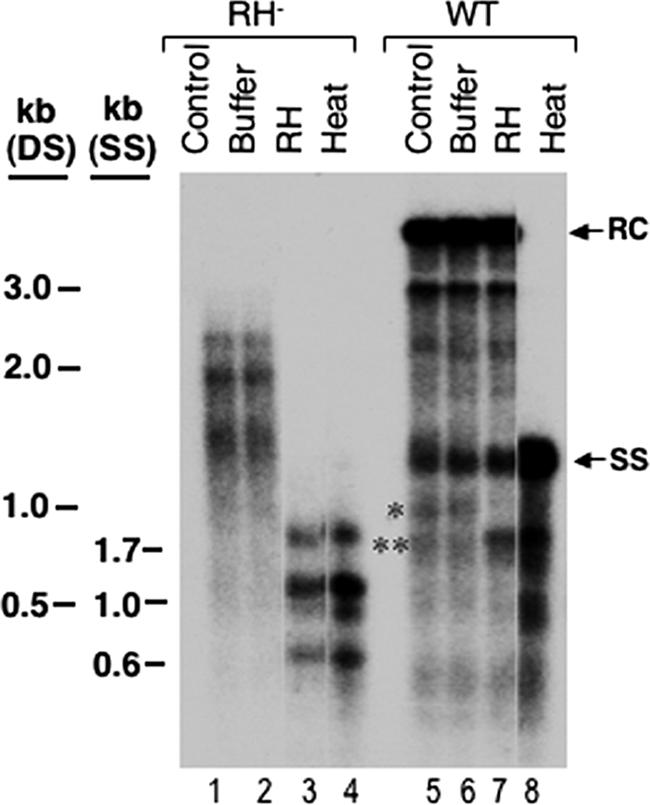
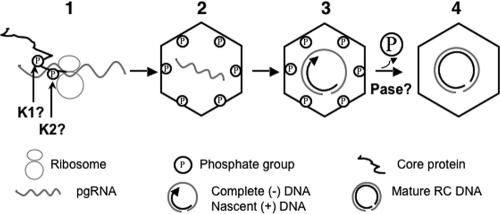
Reverse transcription, an essential step in the life cycle of all retroelements, is a complex, multistep process whose regulation is not yet clearly understood. We have recently shown that reverse transcription in the pararetrovirus duck hepatitis B virus is associated with complete dephosphorylation of the viral core protein, which forms the nucleocapsid wherein reverse transcription takes place. Here we present a genetic study of the role of this dynamic nucleocapsid phosphorylation in regulating viral reverse transcription. Detailed analyses of the reverse transcription products synthesized within nucleocapsids composed of core phosphorylation site mutants revealed that alanine substitutions, mimicking the nonphosphorylated state, completely blocked reverse transcription at a very early stage. In contrast, aspartate substitutions, mimicking the phosphorylated state, allowed complete first-strand DNA synthesis but were severely defective in accumulating mature double-stranded DNA. The latter defect was due to a combination of mutant nucleocapsid instability during maturation and a block in mature second-strand DNA synthesis. Thus, the reversible phosphorylation of the nucleocapsids regulates the ordered progression of reverse transcription.
Figures
References
-
- Bottcher, B., S. A. Wynne, and R. A. Crowther. 1997. Determination of the fold of the core protein of hepatitis B virus by electron cryomicroscopy. Nature 386:88-91. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
