

Edinger-Westphal and pharmacologically stimulated accommodative refractive changes and lens and ciliary process movements in rhesus monkeys
- PMID: 17137577
- PMCID: PMC2901562
- DOI: 10.1016/j.exer.2006.10.003
Edinger-Westphal and pharmacologically stimulated accommodative refractive changes and lens and ciliary process movements in rhesus monkeys
Abstract
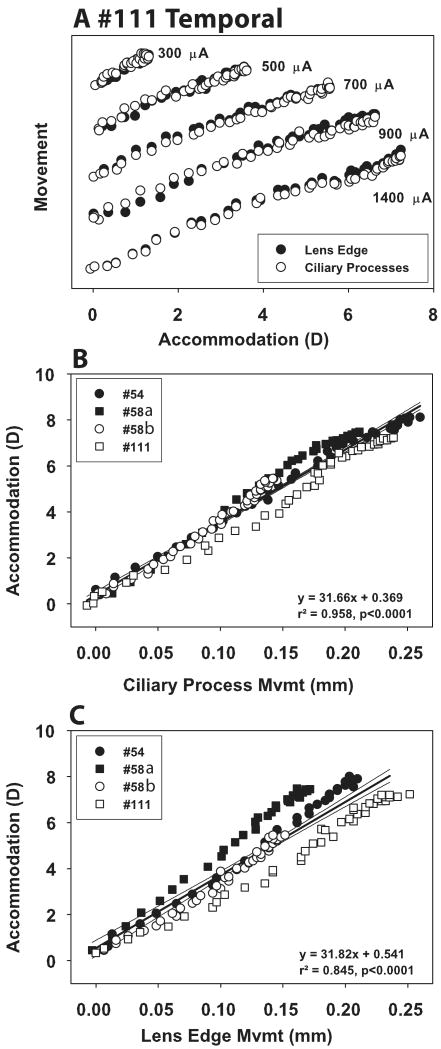
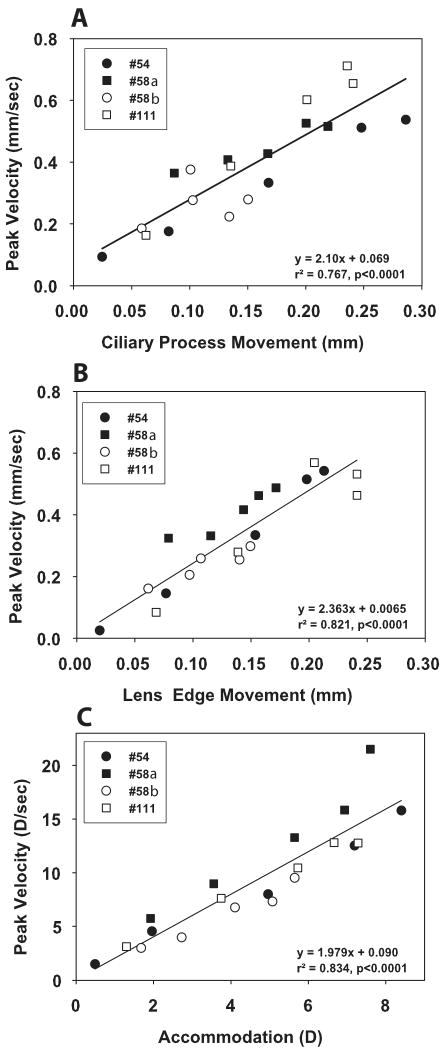
During accommodation, the refractive changes occur when the ciliary muscle contracts, releasing resting zonular tension and allowing the lens capsule to mold the lens into an accommodated form. This results in centripetal movement of the ciliary processes and lens edge. The goal of this study was to understand the relationship between accommodative refractive changes, ciliary process movements and lens edge movements during Edinger-Westphal (EW) and pharmacologically stimulated accommodation in adolescent rhesus monkeys. Experiments were performed on one eye each of three rhesus monkeys with permanent indwelling electrodes in the EW nucleus of the midbrain. EW stimulated accommodative refractive changes were measured with infrared photorefraction, and ciliary process and lens edge movements were measured with slit-lamp goniovideography on the temporal aspect of the eye. Images were recorded on the nasal aspect for one eye during EW stimulation. Image analysis was performed off-line at 30 Hz to determine refractive changes and ciliary body and lens edge movements during EW stimulated accommodation and after carbachol iontophoresis to determine drug induced accommodative movements. Maximum EW stimulated accommodation was 7.36+/-0.49 D and pharmacologically stimulated accommodation was 14.44+/-1.21 D. During EW stimulated accommodation, the ciliary processes and lens edge moved centripetally linearly by 0.030+/-0.001 mm/D and 0.027+/-0.001 mm/D, with a total movement of 0.219+/-0.034 mm and 0.189+/-0.023 mm, respectively. There was no significant nasal/temporal difference in ciliary process or lens edge movements. 30-40 min after pharmacologically stimulated accommodation, the ciliary processes moved centripetally a total of 0.411+/-0.048 mm, or 0.030+/-0.005 mm/D, and the lens edge moved centripetally 0.258+/-0.014 mm, or 0.019+/-0.003 mm/D. The peaks and valleys of the ciliary processes moved by similar amounts during both supramaximal EW and pharmacologically stimulated accommodation. In conclusion, this study shows, for the first time, that the ciliary processes and lens edge move centripetally, linearly with refraction during EW stimulated accommodation. During pharmacological stimulation, the ciliary processes move to a greater extent than the lens edge, confirming that in adolescent monkeys, lens movement limits the accommodative optical change in the eye.
Figures






Comment in
-
Comment on 'Edinger-Westphal and pharmacologically stimulated accommodative refractive changes and lens ciliary process movements in rhesus monkeys' by L.A. Ostrin and A. Glasser [Exp. Eye Res. 84(2) (2007) 302-313].Exp Eye Res. 2007 Aug;85(2):298-9; author reply 300-1. doi: 10.1016/j.exer.2007.04.010. Epub 2007 May 6. Exp Eye Res. 2007. PMID: 17574239 No abstract available.
References
-
- Bito LZ, DeRousseau CJ, Kaufman PL, Bito JW. Age-dependent loss of accommodative amplitude in rhesus monkeys: an animal model for presbyopia. Invest Ophthalmol Vis Sci. 1982;23:23–31. - PubMed
-
- Bito LZ, Kaufman PL, DeRousseau CJ, Koretz J. Presbyopia: an animal model and experimental approaches for the study of the mechanism of accommodation and ocular aging. Eye. 1987;1(Pt 2):222–30. - PubMed
-
- Burd HJ, Judge SJ, Flavell MJ. Mechanics of accommodation of the human eye. Vision Res. 1999;39:1591–5. - PubMed
-
- Coleman DJ, Fish SK. Presbyopia, accommodation, and the mature catenary. Ophthalmology. 2001;108:1544–51. - PubMed
-
- Crawford K, Terasawa E, Kaufman PL. Reproducible stimulation of ciliary muscle contraction in the cynomolgus monkey via a permanent indwelling midbrain electrode. Brain Res. 1989;503:265–72. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
