

Probing the architecture of an L-type calcium channel with a charged phenylalkylamine: evidence for a widely open pore and drug trapping
- PMID: 17138559
- PMCID: PMC3189693
- DOI: 10.1074/jbc.M609153200
Probing the architecture of an L-type calcium channel with a charged phenylalkylamine: evidence for a widely open pore and drug trapping
Abstract
Voltage-gated calcium channels are in a closed conformation at rest and open temporarily when the membrane is depolarized. To gain insight into the molecular architecture of Ca(v)1.2, we probed the closed and open conformations with the charged phenylalkylamine (-)devapamil ((-)qD888). To elucidate the access pathway of (-)D888 to its binding pocket from the intracellular side, we used mutations replacing a highly conserved Ile-781 by threonine/proline in the pore-lining segment IIS6 of Ca(v)1.2 (1). The shifted channel gating of these mutants (by 30-40 mV in the hyperpolarizing direction) enabled us to evoke currents with identical kinetics at different potentials and thus investigate the effect of the membrane potentials on the drug access per se. We show here that under these conditions the development of channel block by (-)qD888 is not affected by the transmembrane voltage. Recovery from block at rest was, however, accelerated at more hyperpolarized voltages. These findings support the conclusion that Ca(v)1.2 must be opening widely to enable free access of the charged (-)D888 molecule to its binding site, whereas drug dissociation from the closed channel conformation is restricted by bulky channel gates. The functional data indicating a location of a trapped (-)D888 molecule close to the central pore region are supported by a homology model illustrating that the closed Ca(v)1.2 is able to accommodate a large cation such as (-)D888.
Figures
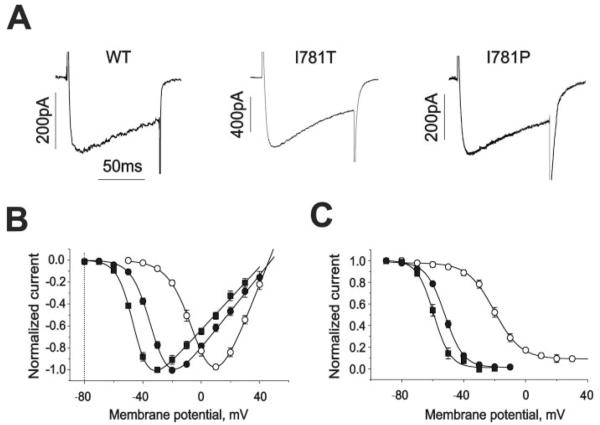
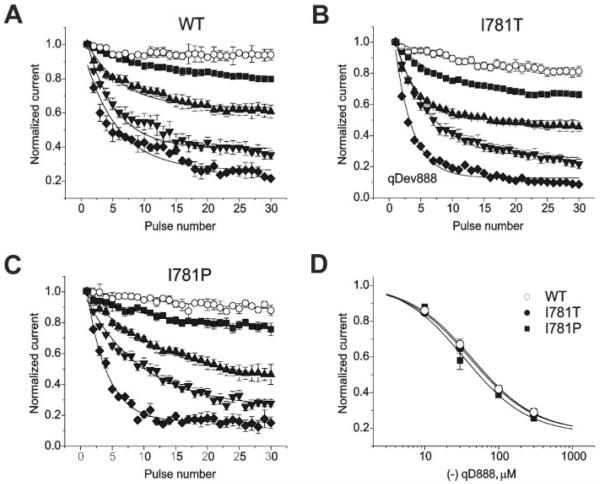
 ) (−)qD888 in the pipette (intracellular) solution. Channel block was estimated as peak IBa inhibition during trains of 30 pulses (0.2 Hz, 100 ms) applied from a holding potential of −80 mV to +20 mV (wild-type), −10 mV (I781T) or −20 mV (I781P). D, concentration dependence of peak IBa inhibition by (−)qD888 in wild-type (○), I781T (●), and I781P (■) mutant channels. Channel block was estimated as the difference between “steady state” normalized current in control and in presence of (−)qD888. Data points are the mean from 3–6 experiments. The IC50 values were obtained by fitting the data points to the Hill equation (as described under “Experimental Procedures”) yielding 43.7 ± 3.4 μ
) (−)qD888 in the pipette (intracellular) solution. Channel block was estimated as peak IBa inhibition during trains of 30 pulses (0.2 Hz, 100 ms) applied from a holding potential of −80 mV to +20 mV (wild-type), −10 mV (I781T) or −20 mV (I781P). D, concentration dependence of peak IBa inhibition by (−)qD888 in wild-type (○), I781T (●), and I781P (■) mutant channels. Channel block was estimated as the difference between “steady state” normalized current in control and in presence of (−)qD888. Data points are the mean from 3–6 experiments. The IC50 values were obtained by fitting the data points to the Hill equation (as described under “Experimental Procedures”) yielding 43.7 ± 3.4 μ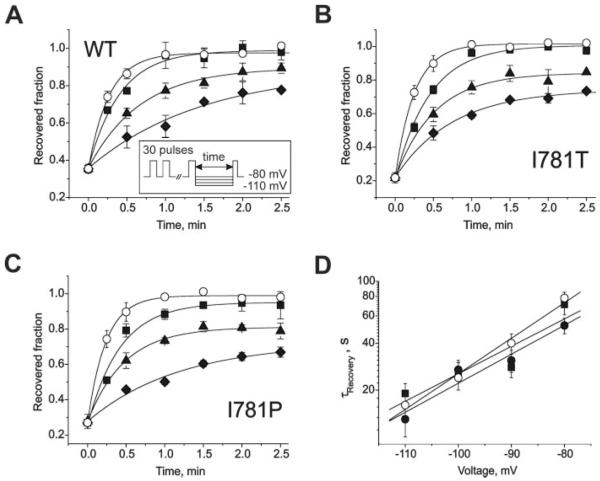 ). Inset, block was elicited by a standard conditioning train of 30 pulses (see also Fig. 2) and recovery measured applying test pulses at different time after the conditioning train. Data points were fitted by mono-exponential functions; yielding time constants: τ−110 = 16 ± 3 s, τ−100 = 24 ± 4 s, τ−90 = 40 ± 6 s, and τ−80 = 78 ± 7 s for wild-type; τ−110 = 13 ± 3 s, τ−100 = 27 ± 4 s, τ−90 = 31 ± 5 s, and τ−80 = 52 ± 6 s for I781T mutant; τ−110 = 19 ± 3 s, τ−100 = 25 ± 5 s, τ−90 = 28 ± 4 s, and τ−80 = 71 ± 10 s for I781P mutant. D, semi-logarithmic plot of the recovery time constants versus holding potentials. Regression lines yield slopes of 0.022 mV−1 (wild-type), 0.018 mV−1 (I781T), and 0.017 mV−1 (I781P).
). Inset, block was elicited by a standard conditioning train of 30 pulses (see also Fig. 2) and recovery measured applying test pulses at different time after the conditioning train. Data points were fitted by mono-exponential functions; yielding time constants: τ−110 = 16 ± 3 s, τ−100 = 24 ± 4 s, τ−90 = 40 ± 6 s, and τ−80 = 78 ± 7 s for wild-type; τ−110 = 13 ± 3 s, τ−100 = 27 ± 4 s, τ−90 = 31 ± 5 s, and τ−80 = 52 ± 6 s for I781T mutant; τ−110 = 19 ± 3 s, τ−100 = 25 ± 5 s, τ−90 = 28 ± 4 s, and τ−80 = 71 ± 10 s for I781P mutant. D, semi-logarithmic plot of the recovery time constants versus holding potentials. Regression lines yield slopes of 0.022 mV−1 (wild-type), 0.018 mV−1 (I781T), and 0.017 mV−1 (I781P).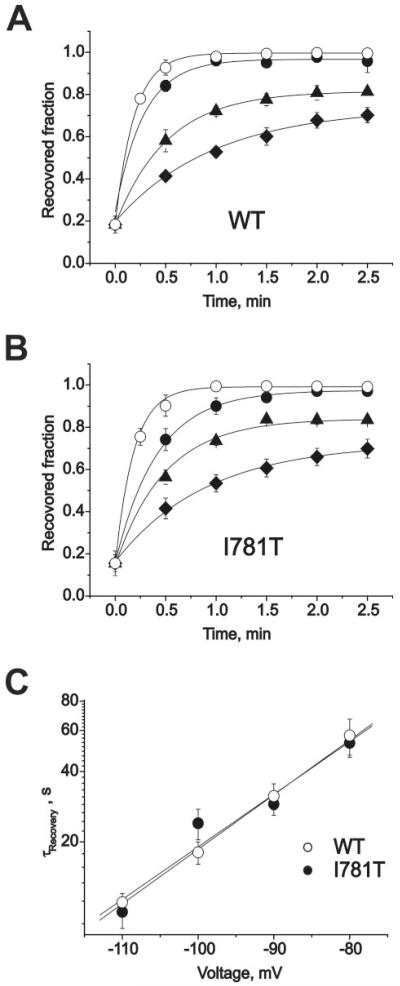 ) (same protocol as described in the legend to Fig. 3). Data points were fitted by mono-exponential functions yielding time constants: τ−110 = 11 ± 1 s,τ−100 = 18 ± 2 s,τ−90 = 31 ± 4 s, and τ−80 = 57 ± 10 s for wild-type; τ−110 = 10 ± 1 s, τ−100 = 24 ± 4 s, τ−90 = 29 ± 3 s, and τ−80 = 53 ± 7 s for I781T mutant. C, semi-logarithmic plot of the recovery time constants versus holding potentials. Regression yields slopes of 0.023 mV−1 (wild-type) and 0.022 mV−1 (I781T).
) (same protocol as described in the legend to Fig. 3). Data points were fitted by mono-exponential functions yielding time constants: τ−110 = 11 ± 1 s,τ−100 = 18 ± 2 s,τ−90 = 31 ± 4 s, and τ−80 = 57 ± 10 s for wild-type; τ−110 = 10 ± 1 s, τ−100 = 24 ± 4 s, τ−90 = 29 ± 3 s, and τ−80 = 53 ± 7 s for I781T mutant. C, semi-logarithmic plot of the recovery time constants versus holding potentials. Regression yields slopes of 0.023 mV−1 (wild-type) and 0.022 mV−1 (I781T).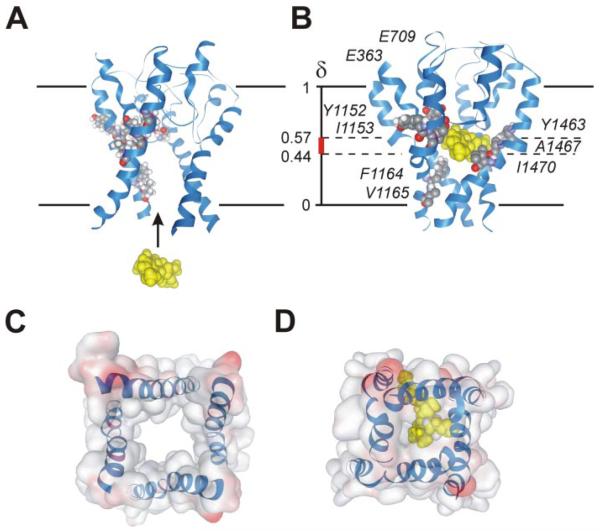
References
-
- Catterall WA. Neuron. 2000;26:13–25. - PubMed
-
- Bezanilla F. Physiol. Rev. 2000;80:555–592. - PubMed
-
- Doyle DA, Morais Cabral J, Pfuetzner RA, Kuo A, Gulbis JM, Cohen SL, Chait BT, MacKinnon R. Science. 1998;280:69–77. - PubMed
-
- Jiang Y, Lee A, Chen J, Cadene M, Chait BT, MacKinnon R. Nature. 2002;417:523–526. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
