

The POLARIS peptide of Arabidopsis regulates auxin transport and root growth via effects on ethylene signaling
- PMID: 17138700
- PMCID: PMC1693943
- DOI: 10.1105/tpc.106.040790
The POLARIS peptide of Arabidopsis regulates auxin transport and root growth via effects on ethylene signaling
Abstract
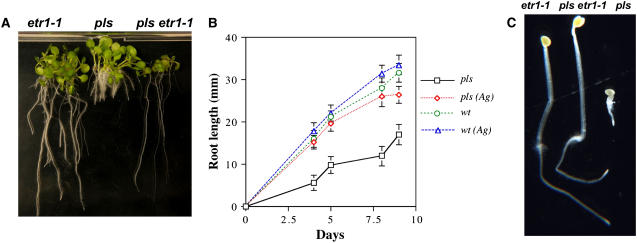
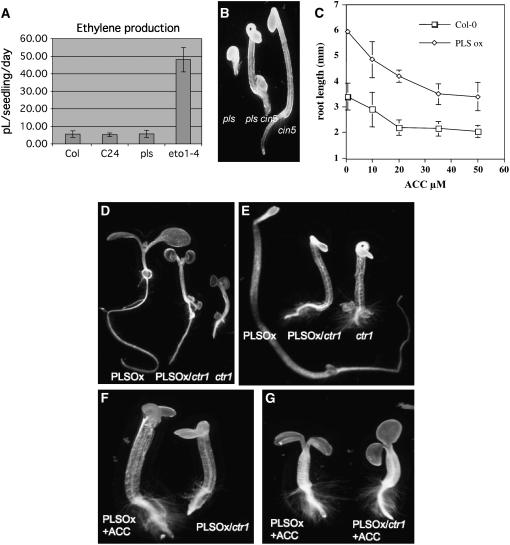
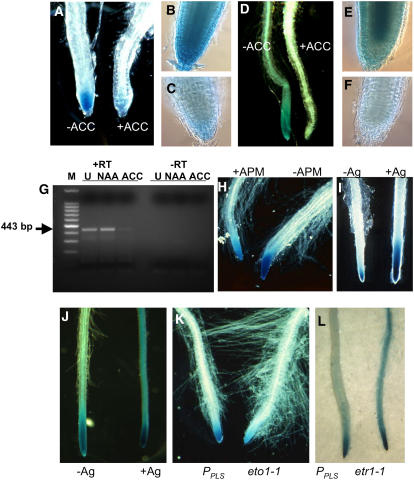
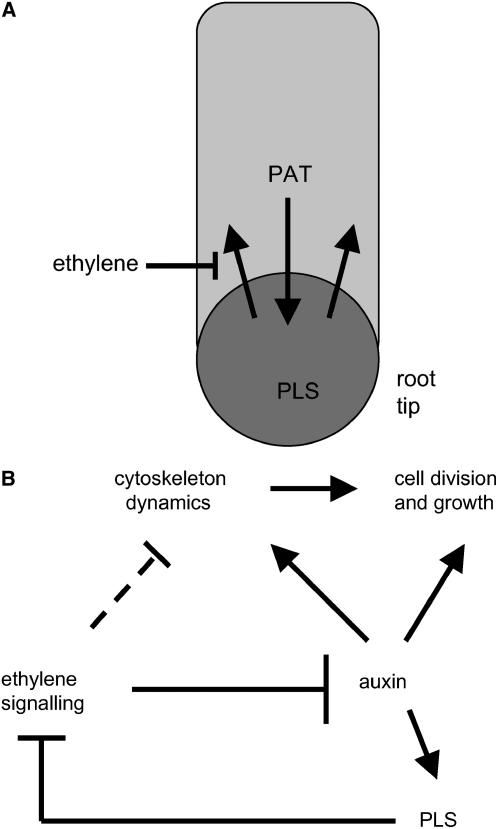
The rate and plane of cell division and anisotropic cell growth are critical for plant development and are regulated by diverse mechanisms involving several hormone signaling pathways. Little is known about peptide signaling in plant growth; however, Arabidopsis thaliana POLARIS (PLS), encoding a 36-amino acid peptide, is required for correct root growth and vascular development. Mutational analysis implicates a role for the peptide in hormone responses, but the basis of PLS action is obscure. Using the Arabidopsis root as a model to study PLS action in plant development, we discovered a link between PLS, ethylene signaling, auxin homeostasis, and microtubule cytoskeleton dynamics. Mutation of PLS results in an enhanced ethylene-response phenotype, defective auxin transport and homeostasis, and altered microtubule sensitivity to inhibitors. These defects, along with the short-root phenotype, are suppressed by genetic and pharmacological inhibition of ethylene action. PLS expression is repressed by ethylene and induced by auxin. Our results suggest a mechanism whereby PLS negatively regulates ethylene responses to modulate cell division and expansion via downstream effects on microtubule cytoskeleton dynamics and auxin signaling, thereby influencing root growth and lateral root development. This mechanism involves a regulatory loop of auxin-ethylene interactions.
Figures




References
-
- Abe, T., Thitamadee, S., and Hashimoto, T. (2004). Microtubule defects and cell morphogenesis in the lefty1lefty2 tubulin mutant of Arabidopsis thaliana. Plant Cell Physiol. 45 211–220. - PubMed
-
- Abeles, F., Morgan, P., and Saltveit, M. (1992). Ethylene in Plant Biology. San Diego, CA: Academic Press.
-
- Anthony, R.G., and Hussey, P.J. (1999. a). Dinitroaniline herbicide resistance and the microtubule cytoskeleton. Trends Plant Sci. 4 112–116. - PubMed
-
- Anthony, R.G., and Hussey, P.J. (1999. b). Double mutation in Eleusine indica alpha-tubulin increases the resistance of transgenic maize calli to dinitroaniline and phosphorothioamidate herbicides. Plant J. 18 669–674. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
