

NoRC-dependent nucleosome positioning silences rRNA genes
- PMID: 17139253
- PMCID: PMC1698900
- DOI: 10.1038/sj.emboj.7601454
NoRC-dependent nucleosome positioning silences rRNA genes
Abstract
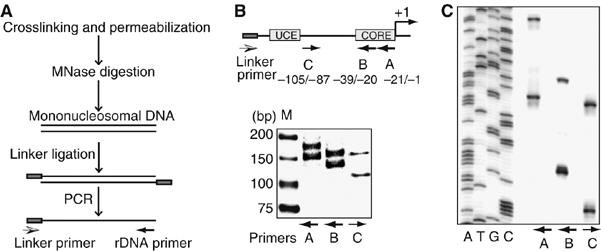
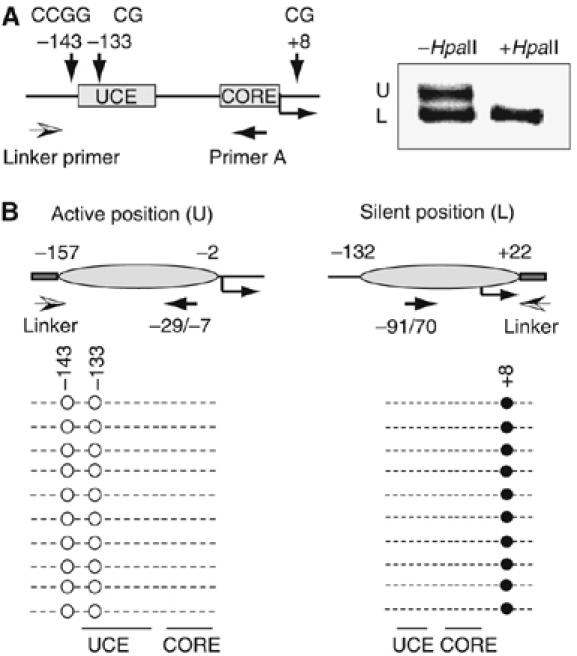
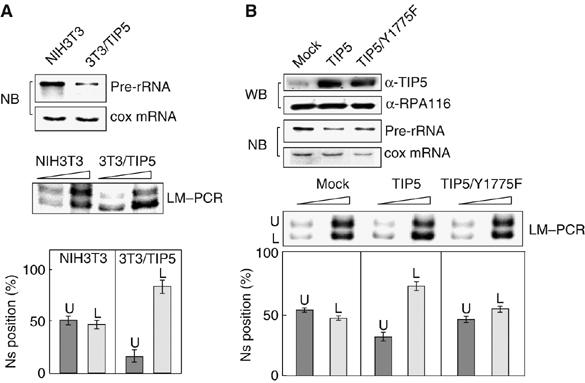
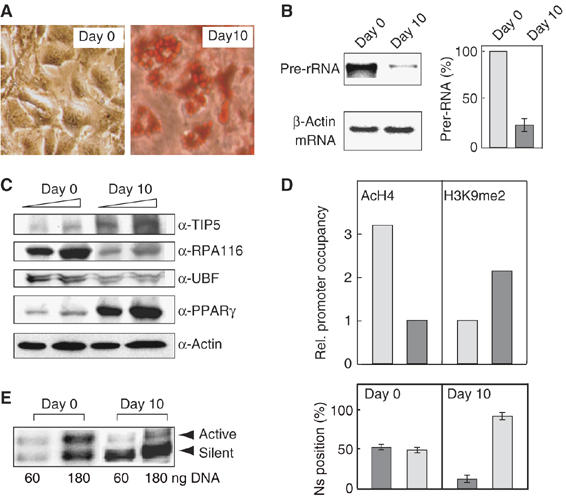
Previous studies have established that the Snf2h-containing chromatin remodeling complex NoRC mediates epigenetic silencing of a subset of rRNA genes (rDNA) by recruiting enzymatic activities that modify histones and methylate DNA. Here we have analyzed nucleosome positions at the murine rDNA promoter and show that active and silent rDNA copies are characterized not only by specific epigenetic marks but also by differently positioned nucleosomes. At active genes the promoter-bound nucleosome covers nucleotides from -157 to -2, whereas at silent genes the nucleosome is positioned 25 nucleotides further downstream. We provide evidence that NoRC is the molecular machine that shifts the promoter-bound nucleosome downstream of the transcription start site into a translational position that is unfavorable for transcription complex formation.
Figures
References
-
- Bazett-Jones DP, Leblanc B, Herfort M, Moss T (1994) Short-range DNA looping by the Xenopus HMG-box transcription factor, xUBF. Science 264: 1134–1137 - PubMed
-
- Cooper JP, Roth SY, Simpson RT (1994) The global transcriptional regulators, SSN6 and TUP1, play distinct roles in the establishment of a repressive chromatin structure. Genes Dev 8: 1400–1410 - PubMed
-
- Corona DF, Tamkun JW (2004) Multiple roles for ISWI in transcription, chromosome organization and DNA replication. Biochim Biophys Acta 1677: 113–119 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
