

Genome-wide natural antisense transcription: coupling its regulation to its different regulatory mechanisms
- PMID: 17139297
- PMCID: PMC1794690
- DOI: 10.1038/sj.embor.7400857
Genome-wide natural antisense transcription: coupling its regulation to its different regulatory mechanisms
Abstract
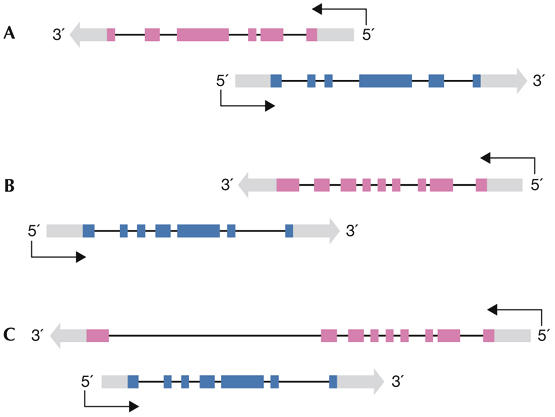
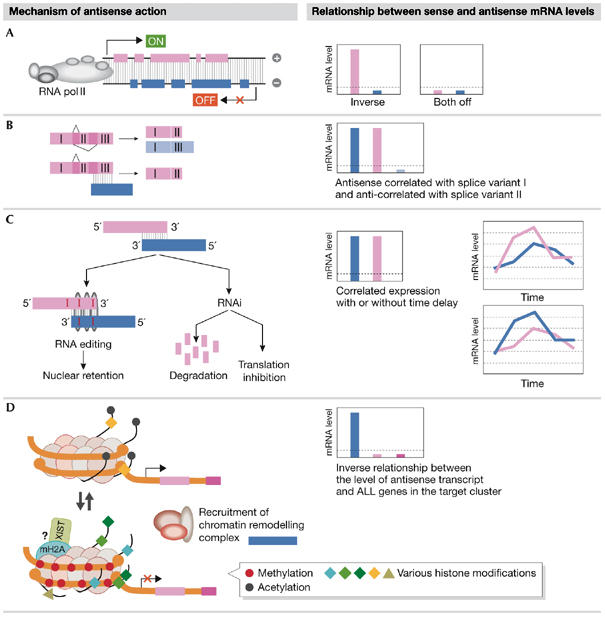
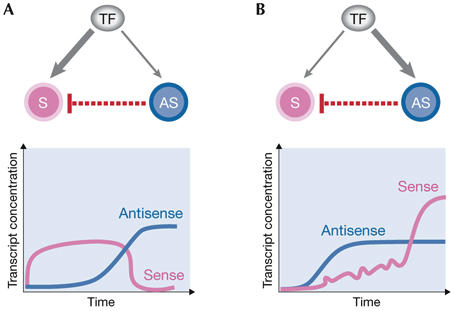
Many genomic loci contain transcription units on both strands, therefore two oppositely oriented transcripts can overlap. Often, one strand codes for a protein, whereas the transcript from the other strand is non-encoding. Such natural antisense transcripts (NATs) can negatively regulate the conjugated sense transcript. NATs are highly prevalent in a wide range of species--for example, around 15% of human protein-encoding genes have an associated NAT. The regulatory mechanisms by which NATs act are diverse, as are the means to control their expression. Here, we review the current understanding of NAT function and its mechanistic basis, which has been gathered from both individual gene cases and genome-wide studies. In parallel, we survey findings about the regulation of NAT transcription. Finally, we hypothesize that the regulation of antisense transcription might be tailored to its mode of action. According to this model, the observed relationship between the expression patterns of NATs and their targets might indicate the regulatory mechanism that is in action.
Figures


References
-
- Aravin AA, Naumova NM, Tulin AV, Vagin VV, Rozovsky YM, Gvozdev VA (2001) Double-stranded RNA-mediated silencing of genomic tandem repeats and transposable elements in the D. melanogaster germline. Curr Biol 11: 1017–1027 - PubMed
-
- Barrell BG, Air GM, Hutchison CA 3rd (1976) Overlapping genes in bacteriophage φX174. Nature 264: 34–41 - PubMed
-
- Bertone P et al. (2004) Global identification of human transcribed sequences with genome tiling arrays. Science 306: 2242–2246 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
