

Metalloprotein and metallo-DNA/RNAzyme design: current approaches, success measures, and future challenges
- PMID: 17140190
- PMCID: PMC2533576
- DOI: 10.1021/ic052007t
Metalloprotein and metallo-DNA/RNAzyme design: current approaches, success measures, and future challenges
Abstract
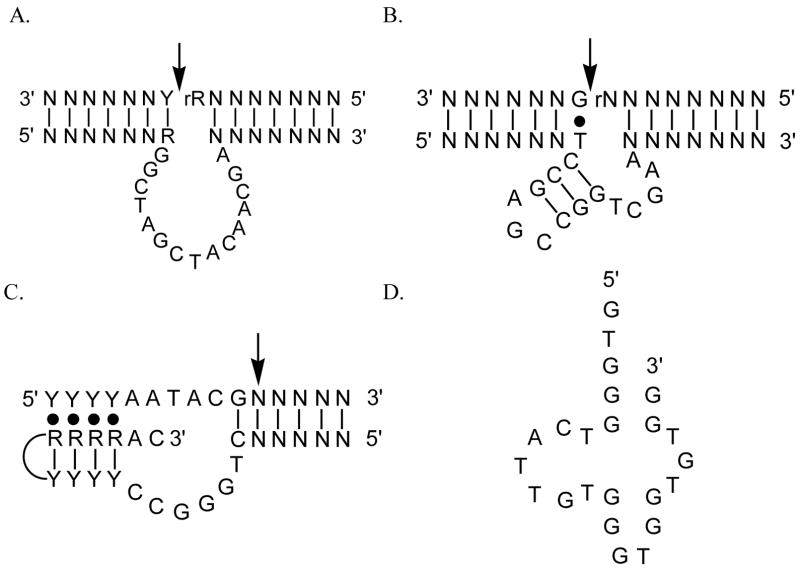
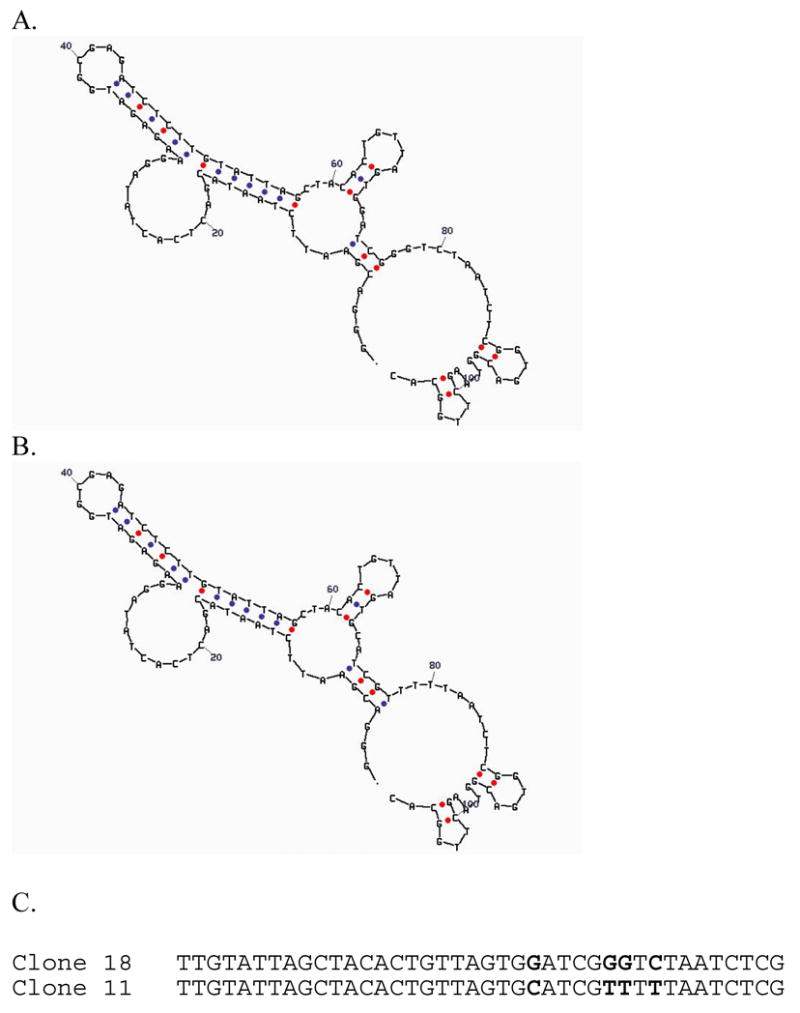
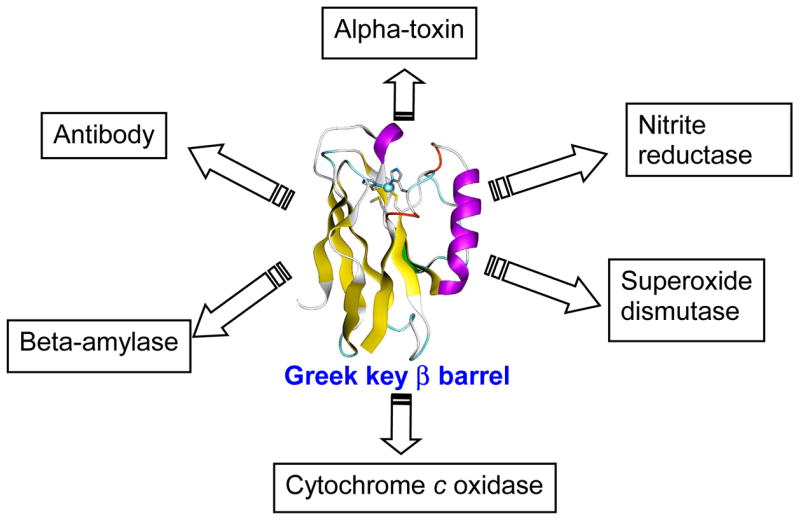
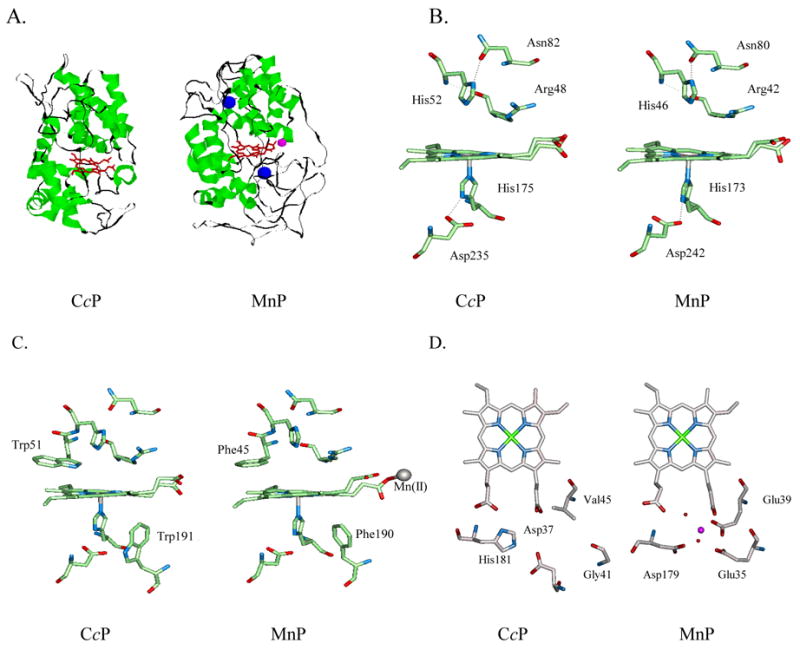
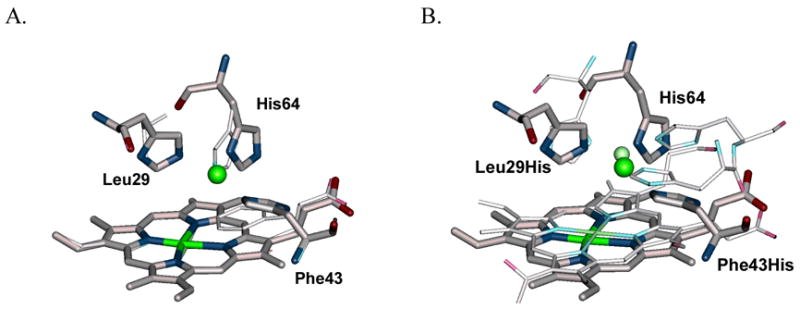
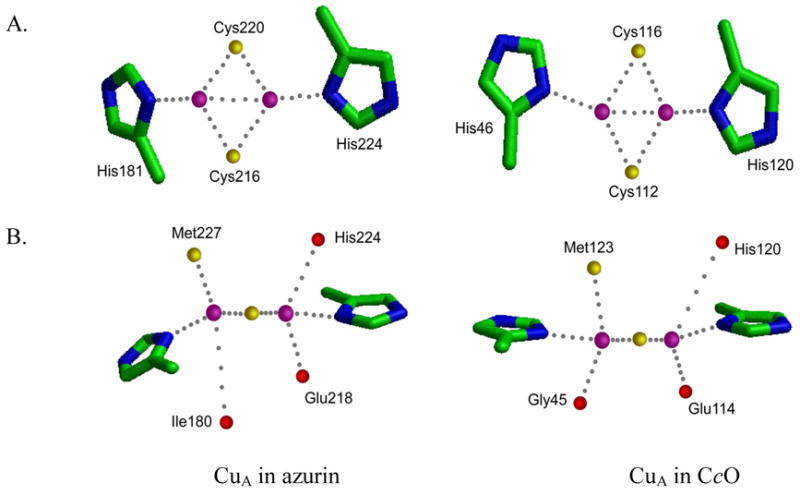
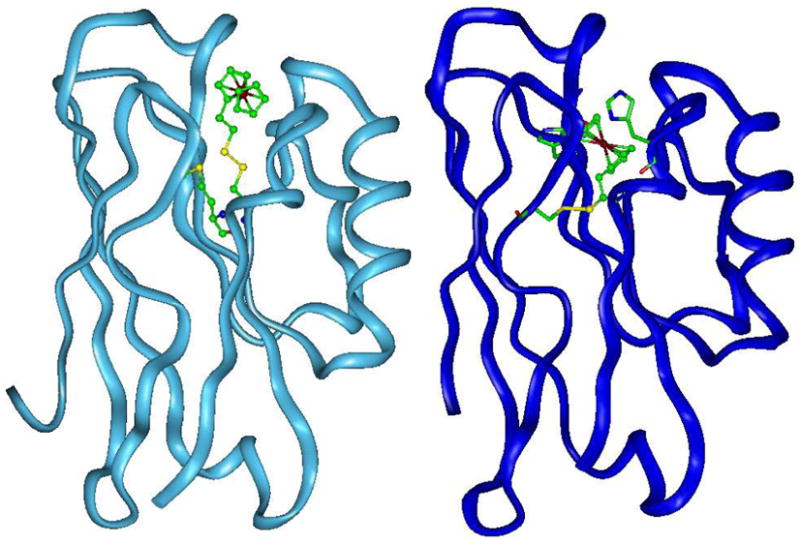
Specific metal-binding sites have been found in not only proteins but also DNA and RNA molecules. Together these metalloenzymes consist of a major portion of the enzyme family and can catalyze some of the most difficult biological reactions. Designing these metalloenzymes can be both challenging and rewarding because it can provide deeper insights into the structure and function of proteins and cheaper and more stable alternatives for biochemical and biotechnological applications. Toward this goal, both rational and combinatorial approaches have been used. The rational approach is good for designing metalloenzymes that are well characterized, such as heme proteins, while the combinatorial approach is better at designing those whose structures are poorly understood, such as metallo-DNA/RNAzymes. Among the rational approaches, de novo design is at its best when metal-binding sites reside in a scaffold whose structure has been designed de novo (e.g., alpha-helical bundles). Otherwise, design using native scaffolds can be equally effective, allowing more choices of scaffolds whose structural stability is often more resistant to multiple mutations. In addition, computational and empirical designs have both enjoyed successes. Because of the limitation in defining structural parameters for metal-binding sites, a computational approach is restricted to mostly metal-binding sites that are well defined, such as mono- or homonuclear centers. An empirical approach, even though it is less restrictive in the metal-binding sites to be designed, depends heavily on one's knowledge and choice of templates and targets. An emerging approach is a combination of both computational and empirical approaches. The success of these approaches can be measured not only by three-dimensional structural comparison between the designed and target enzymes but also by the total amount of insight obtained from the design process and studies of the designed enzymes. One of the biggest advantages of designed metalloenzymes is the potential of placing two different metal-binding sites in the same protein framework for comparison. A final measure of success is how one can utilize the insight gained from the intellectual exercise to design new metalloenzymes, including those with unprecedented structures and functions. Future challenges include designing more complex metalloenzymes such as heteronuclear metal centers with strong nanomolar or better affinities. A key to meeting this challenge is to focus on the design of not only primary but also secondary coordination spheres using a combination of improved computer programs, experimental design, and high-resolution crystallography.
Figures




References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
