

Transcriptional regulation by competing transcription factor modules
- PMID: 17140283
- PMCID: PMC1676028
- DOI: 10.1371/journal.pcbi.0020164
Transcriptional regulation by competing transcription factor modules
Abstract
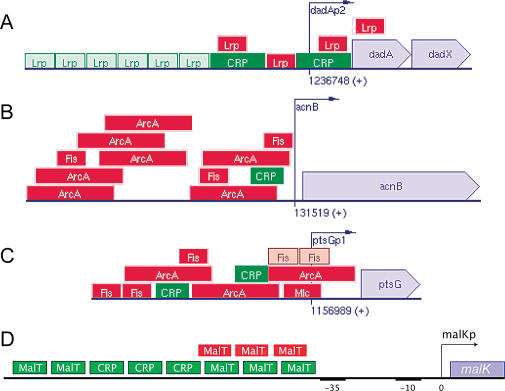
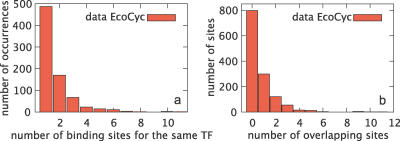
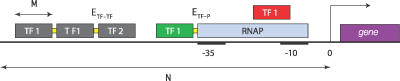
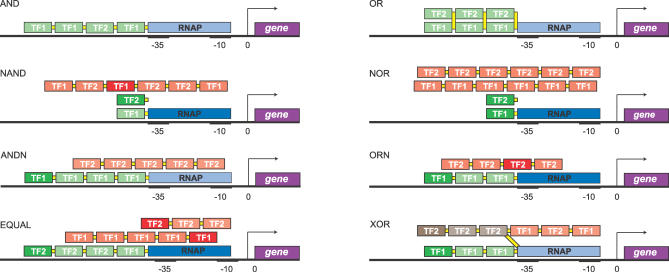
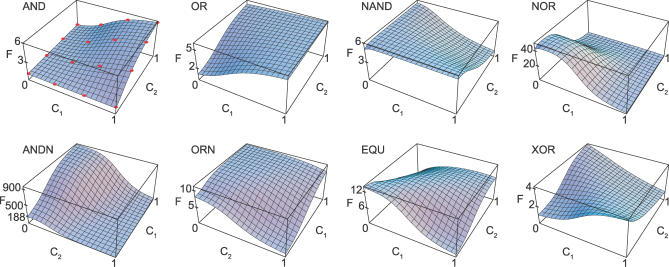
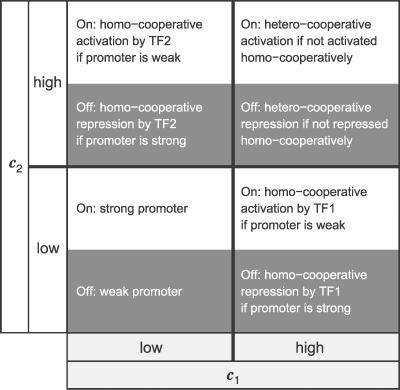
Gene regulatory networks lie at the heart of cellular computation. In these networks, intracellular and extracellular signals are integrated by transcription factors, which control the expression of transcription units by binding to cis-regulatory regions on the DNA. The designs of both eukaryotic and prokaryotic cis-regulatory regions are usually highly complex. They frequently consist of both repetitive and overlapping transcription factor binding sites. To unravel the design principles of these promoter architectures, we have designed in silico prokaryotic transcriptional logic gates with predefined input-output relations using an evolutionary algorithm. The resulting cis-regulatory designs are often composed of modules that consist of tandem arrays of binding sites to which the transcription factors bind cooperatively. Moreover, these modules often overlap with each other, leading to competition between them. Our analysis thus identifies a new signal integration motif that is based upon the interplay between intramodular cooperativity and intermodular competition. We show that this signal integration mechanism drastically enhances the capacity of cis-regulatory domains to integrate signals. Our results provide a possible explanation for the complexity of promoter architectures and could be used for the rational design of synthetic gene circuits.
Conflict of interest statement
Figures
References
-
- Jacob F, Monod J. Genetic regulatory mechanisms in the synthesis of proteins. J Mol Biol. 1961;3:318–356. - PubMed
-
- Müller-Hill B. The lac operon: A short history of a genetic paradigm. Berlin: Walter de Gruyter; 1996. 207
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
