

Sound sequence discrimination learning motivated by reward requires dopaminergic D2 receptor activation in the rat auditory cortex
- PMID: 17142301
- PMCID: PMC1783622
- DOI: 10.1101/lm.390506
Sound sequence discrimination learning motivated by reward requires dopaminergic D2 receptor activation in the rat auditory cortex
Abstract
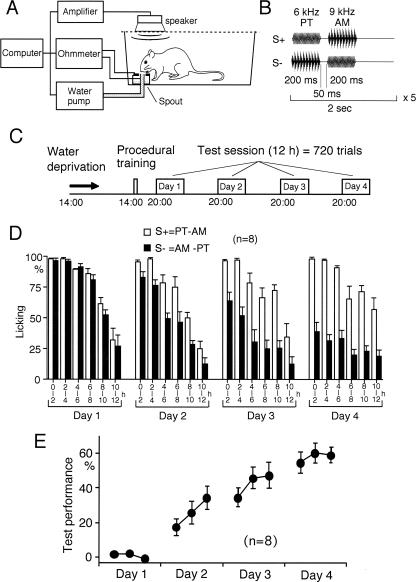
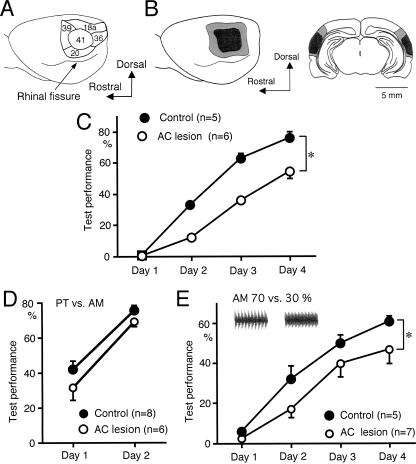
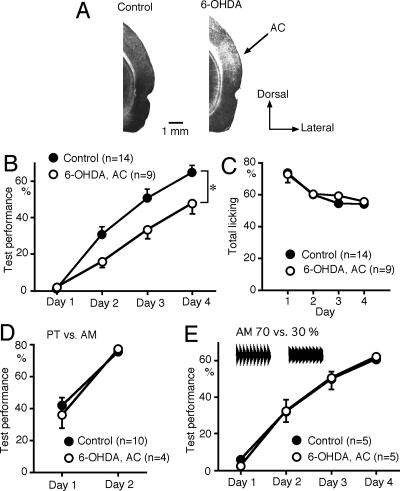
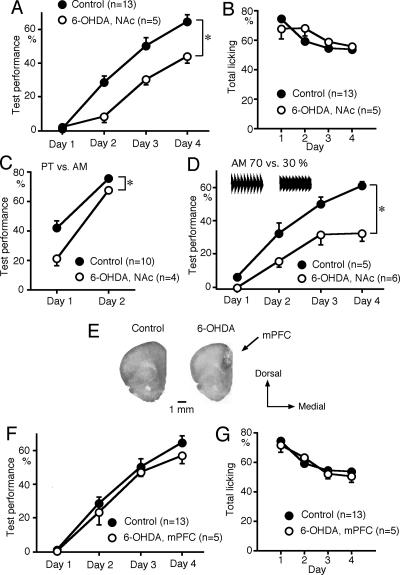
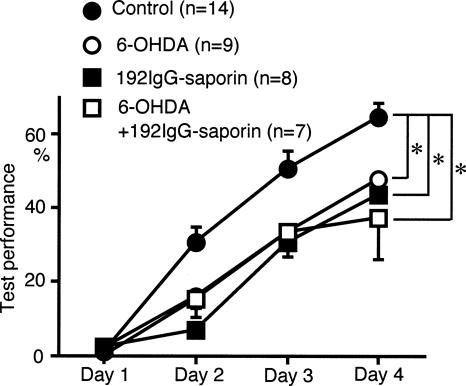
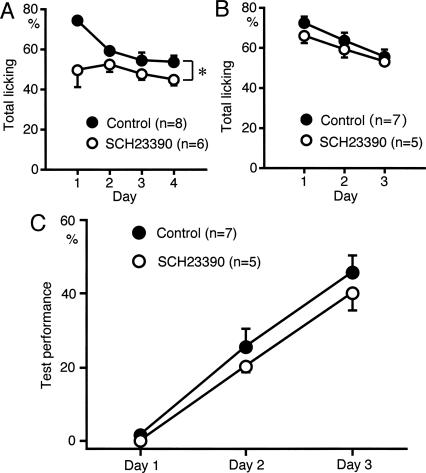
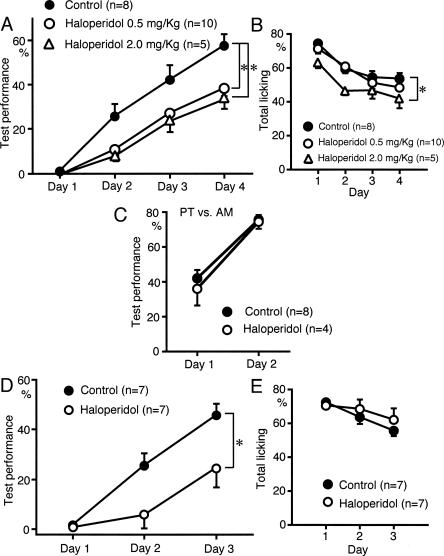
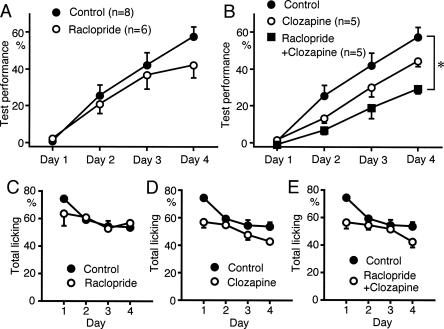
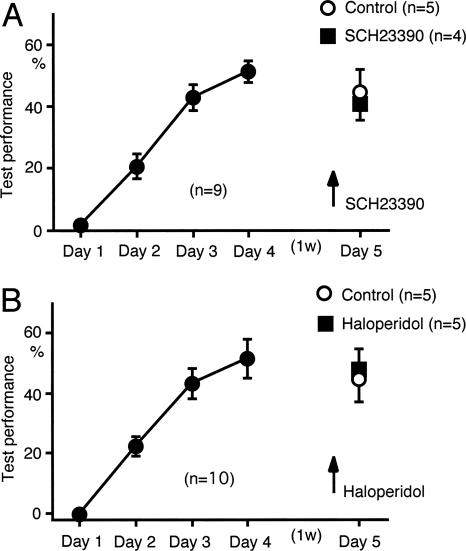
We have previously reported that sound sequence discrimination learning requires cholinergic inputs to the auditory cortex (AC) in rats. In that study, reward was used for motivating discrimination behavior in rats. Therefore, dopaminergic inputs mediating reward signals may have an important role in the learning. We tested the possibility in the present study. Rats were trained to discriminate sequences of two sound components, and licking behavior in response to one of the two sequences was rewarded with water. To identify the dopaminergic inputs responsible for the learning, dopaminergic afferents to the AC were lesioned with local injection of 6-hydroxydopamine (6-OHDA). The injection attenuated sound sequence discrimination learning, while it had no effect on discrimination between the sound components of the sequence stimuli. Local injection of 6-OHDA into the nucleus accumbens attenuated sound discrimination learning. However, not only discrimination learning of sound sequence but also that of the sound components were impaired. SCH23390 (0.2 mg/kg, i.p.), a D1 receptor antagonist, had no effect on sound sequence discrimination learning, while it attenuated the licking behavior to unfamiliar stimuli. Haloperidol (0.5 mg/kg, i.p.), a D2 family antagonist, attenuated sound sequence discrimination learning, while it had no clear suppressive effect on discrimination of two different sound components and licking. These results suggest that D2 family receptors activated by dopaminergic inputs to the AC are required for sound sequence discrimination learning.
Figures
References
-
- Amalric M., Koob G.F. Functionally selective neurochemical afferents and efferents of the mesocorticolimbic and nigrostriatal dopamine system. Prog. Brain Res. 1993;99:209–226. - PubMed
-
- Bao S., Chan V.T., Merzenich M.M. Cortical remodelling induced by activity of ventral tegmental dopamine neurons. Nature. 2001;412:79–83. - PubMed
-
- Barnett A., Iorio L.C., McQuade R., Chipkin R.E. Pharmacological and behavioral effects of D1 dopamine antagonists. Adv. Exp. Med. Biol. 1988;235:137–144. - PubMed
-
- Blond O., Crepel F., Otani S. Long-term potentiation in rat prefrontal slices facilitated by phased application of dopamine. Eur. J. Pharmacol. 2002;438:115–116. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases